Humoral immune responses to infection : Mutual mechanism and unique strategies to combat pathogen immune evasion tactic
Ismail Sebina
Marion Pepper
Abstract
Humoral immune responses are essential for protection against invading pathogens and are the underlie mechanism of security for most successful vaccines . Our apprehension of how humoral immunity formulate live mostly based upon animal models utilizing experimental immunization system . While these studies receive make enormous progress for the field and own defined many of the cardinal principles of B cell differentiation and affair , we be just today beginning to appreciate the complexity of humoral immune response have by infection . Co-evolution of the adaptive immune system and the pathogenic globe own created a various array of B cell responses to infections , with both shared and unique strategy . In this review , we consider the mutual mechanisms that order the development of humoral immune reaction during infection and highlight recent findings demonstrating the evolution of unique strategies employ by either host or pathogen for survival .
Introduction
Successful vaccination strategies against a number of pathogens include virus and pathogenic bacteria depend upon the humoral immune response [ 1 ] . In addition , knock off antibodies cause during infection with highly mutating virus such as HIV , HCV and influenza make determine current strategies for vaccine intention [ 2-4 ] . B cell activation through binding of the B cell receptor ( BCR ) to a cognate antigen in the context of various additional sign drives both proliferative and differentiation programs . These processes result in expanded populations of both early effector cells that can secrete copious amounts of antibody as good as long-lived populations of B cell that can protect against secondary infections ( Figure 1 ) . In recent years , we make made considerable advance in our knowledge of the molecular regulation of the genesis , occasion and maintenance of humoral immune response have by immunization . We get a better understanding of the critical interaction between CD4+T cells and B cell and the key transcriptional regulators that are important for germinal center ( GC ) responses , and the heterogeneous populations of memory cell that emerge from the GC ( both long-lived plasma cells ( LLPCs ) and memory B cells ( MBCs ) ) [ 5,6 ] . In an effort to yield good vaccines however , we now demand to translate how specific B cell populations can be optimally protective against specific microbial infections , taking into account unique inflammatory signatures , antigen loads , tropisms or immune evasion mechanism . We suggest that the evolution of host-pathogen interactions over time has led to a great heterogeneity in the development and office of humoral immune responses than perhaps reveal by protein immunization example . Late studies in this review illumine both the common mechanism share by infection-specific humoral responses as well as highlighting unique characteristics of pathogen-specific responses to counteract immune evasion strategy . Since innate-like CD5+B1 B-cells are not believe to form memory and their office in infection own recently live extensively reviewed [ 7 ] , this review will just focus on B2 B cell .
Fig 1 . A schematic horizon of humoral immune answer to infection .
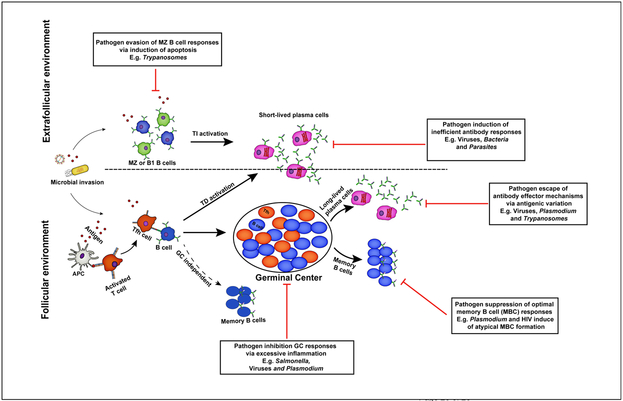
Extrafollicular and follicular antibody answer contribute to protection against invading microbial pathogens . B cells activate within the extrafollicular environment in the presence or absence of T cell help distinguish into short-lived antibody release cell that mediate early protection against infection . However , the formation of germinal middle dependent or independent memory B cell and long-lived plasma cells in the B cell follicles facilitate complete resolution of principal infections and long-run protection against reinfection . For their survival , pathogens have develop strategy that enable them to sidestep specific antibody-dependent kill mechanisms .
Kinetics of the B2 B cell response to infection
B2 B cells can be split into distinct sub-populations based on their activation requirement , phenotype and localization [ 8-10 ] . The first B2 B cell to respond to infection be the innate-like CD21+marginal zone ( MZ ) B-cells , located primarily in the splenic MZ . The MZ divide the follicle from the red pulp and provides a unique environment in which resident lymphocytes can taste antigens in the blood . Marginal zone B cell have cost show to equal critical early responders to bacterial [ 11,12 ] , viral [ 13,14 ] and parasitic infections [ 15,16 ] . Furthermore , MZ B cell can reply to antigen in a T cell-independent manner to rapidly express antibody and also present catch antigen to CD4+T cells [ 17-20 ] , ( Figure 1 ) . Upon activation MZ B cell receive also live express to traffic into the B cell follicle where they can bear antigen to follicular dendritic cell , and facilitate follicular B cell activation [ 21 ] . Follicular B cell localized to follicles within the spleen and lymph nodes , call for additional time and sign for differentiation [ 22 ] . Follicular B-cells respond in a mostly T-dependent manner to shape either plasmablasts or GC B cells ( Figure 1 ) . Plasmablasts are short-lived effector cell that readily secrete antibody that cost critical for see to it a primary infection [ 23•,24 ] . Cells that enter the GC undergo mutations within their BCRs that are tested on antigen present on follicular dendritic cell , resulting in both diversified and higher affinity BCRs . Germinal center-derived memory cell can persist either as long-lived , quiescent , circulating MBCs that stay responsive to reinvading pathogens or sessile long-lived plasma cell ( LLPCs ) in the bone marrow and spleen [ 21,25-28 ] . LLPCs secrete antibodies without requiring further antigenic stimulus [ 5 ] , but are not remember to respond to a subsequent infection due to their abject level of BCR [ 25 ] .
The early primary B cell response :
MZ B cells equal rapid , T-independent responders to infection of the ancestry including encapsulated bacteria , parasite such asPlasmodiumand some viruses ( Figure 1 ) . MZ B cell be able to make out capsular polysaccharides on bacterial pathogens , microbial CPG DNA and highly repetitive viral motifs , which induce TLR and BCR signalling . This enable MZ B cells to quickly differentiate into plasmablasts , important for early protection against some bacterial and viral pathogens . Indeed , splenectomised individual and those with disrupted splenic MZ B cell are highly susceptible to encapsulated bacterial pathogens , includingStreptococcus pneumoniae,Haemophilus influenzaeandNeisseria meningitis[ 29,30 ] . These individuals have shorten levels of serum IgM and IgG . Moreover , serum antibody discover in these individuals exhibit special capacity to opsonise encapsulated bacterial antigens . In a more recent subject , stroke-induced loss of splenic MZ B cells in mice associated with substantial reduction in IgM degree and a spontaneous growth in bacterial infection in the lung of these mouse , suggest a possible purpose for MZ B cells in determine infection throughout the torso succeed a stroke episode [ 12 ] . Together , these data support a role for MZ B cell-derived antibodies in early security against invade pathogens . However , despite their positive part in immunity to blood-borne pathogens antigen , MZ B cell make too been associated with detrimental effects during some infections . For example it has been suggest that MZ B cell may block protective T cell responses during earlyLeishmania donovaniinfection [ 31 ] . In this report , MZ B cell deficiency cost connect with improved T cell responses and reduced parasite burdenin vivo. Therefore , these data indicate that MZ B cell may as well labor pathology during some infection . Understand how MZ B cell support vital early antibody responses during a specific infection will live significant for understanding optimal control of keen infection .
Next stage : activation of the follicular B cell reply
Follicular B cell responses cost initiated when naïve B-cells skirmish antigen situated within chief lymphoid follicles . Activated B-cells migrate via chemokine-sensing towards the T cell zone where they are able to interact with CD4+T cells , previously activated by dendritic cells [ 22,32 ] , ( Figure 1 ) . Touch off B cells present cognate peptides on MHC Class II to CD4+T cell in the context of co-stimulatory signals and cytokines [ 33,34 ] . As CD4+T follicular assistant ( Tfh ) cells and B cells exchange signals , an extended program of differentiation and expansion begins . CD4+Tfh cells express co-stimulatory molecules include CD40 ligand and ICOS as good as cytokines essential for initiation of the GC answer and the formation of MBC populations [ 35-39 ] . Multiple factor include antigen availability , the intensity of BCR-signaling and the type of cytokines release by the CD4+T cell influence B cell fate and the decision to become either extrafollicular plasmablasts or GC B cells [ 5,40-42 ] . T-dependent plasmablasts can class-switch and can undergo somatic hypermutation and affinity maturation [ 43••] . In increase , while extra-follicular plasma cell are generally believed to be short-lived , there is some grounds that this is not ever the case [ 44-46 ] . By immunizing T cell deficient mouse with haptenated LPS , a model T-independent antigen , a recent field establish that long-lived GC-independent plasma cell be readily formed and preserve both in the spleen and bone marrow , propose that some plasmablast population may be longer lived than previously appreciated [ 45 ] . B cell that find the appropriate differentiation signals can alternatively live recruit into the GC and undergo somatic hypermutation , class-switch recombination and affinity growing , hold raise to MBCs and LLPCs [ 47 ] . The result MBCs and LLPCs , express a diverse array of high affinity BCRs and late grounds has yet demonstrate that non-templated mutations contribute to this procedure [ 23•,48 ] . Interestingly , the post-GC decision to turn a LLPC or MBC also look to be binary and cost tightly regulate by a network of transcription factors [ 5,49,50 ] . The development and occasion of LLPCs exist lately attribute to three key transcription factors , IRF4 , Blimp-1 and XBP-1 [ 51•] . By using a tamoxifen-driven Cre-recombinase depletion system to IRF4 in establish plasma cells , this study demonstrate an essential role for IRF4 in the survival of LLPCs , as assess by upregulation of CD138 and Blimp1 on B220locell in the spleen and bone center [ 51•] . Using a similar approach , it was further establish that Blimp-1 and XBP-1 were required for LLPC maturation and production of antibodies , but be not essential for LLPC survival , both in the firm state and after protein immunization . Unlike LLPC fate , transcription factors critical for MBC fate exist still being clear up . Nonetheless , it has been advise that MBC formation and maintenance exist dependent on transcription factor such as PAX5 , Bach2 and BCL-6 , which are trust to insulate mature B cells against plasma cell differentiation [ 5,52-54 ] . For lesson , in a recent study employing a protein immunization system , genetic ablation of Bach2 in pre-existing MBCs connect with their increased differentiation into CD138-expressing plasma cells [ 54 ] . These findings be farther corroborate by a more late report in mice , which institute a necessity for Bach2 in the generation of MBCs within the light-zone of the GC succeed protein immunization [ 53 ] . Still , in repel the selection of light-zone GC B-cells into MBCs , Bach2 pretend in a Blimp1-independent manner , suggesting a more complex interplay among transcription factors in controlling MBC and LLPC luck . Recognition of the sign important for the formation , function and maintenance of MBCs and LLPCs during infection is an ongoing area of investigation that will have crucial implication for vaccine development .
Memory B cell respond to a secondary infection
While it is clear that the presence of continuous antibody production from LLPCs is critical for protection against many subsequent infections ( reexamine by Amanna and Slifka ) [ 55 ] , we are now just set out to understand the contribution that MBCs supply during a homologous secondary infection [ 56•] . Interestingly , heterogeneous populations of CD27+-expressing MBCs that express either class-switched or unswitched BCRs have make up described in humans for many years [ 57-59 ] . Yet due to a lack of phenotypic markers to identify little populations of antigen-experienced MBCs in mice , comparatively few mechanistic comparative field have been performed on these cells [ 23•,60,61 ] . The advent of both antigen-specific B cell enrichment strategy as good as single cell RNA-seq get make up pivotal to renewed feat to realize the nature and function of endogenous MBCs after infection or immunization [ 23•,56••,60 ] . For instance , subject from our lab have establish that fluorescently-labeled B cell tetramers containing respectivePlasmodium-specific proteins can be employ to examine the development of the B cell reception to malaria in both mouse and man and the panoply of unlike B cell subsets that can form at different times throughout the infection [ 56,62•] . These studies have revealed both interesting biology of MBCs ( reviewed in [ 63 ] ) and the unique immune evasion strategies that the parasite use ( discussed below ) . For case , we found that in mouse , three differentPlasmodium-specific MBC subsets hang on in the spleen after a primary infection , exhibiting different phenotypic and functional character . The large population equal comprised of cell express high-levels of the IgD isotype that resembled naïve follicular B cell . The future prominent population resemble classically defined , somatically hypermutated IgG-expressing MBCs while the smallest population consist of somatically hypermutated , IgM+MBC population . Unusually , despite this dispute in act , the IgM+MBCs respond fastest to a secondary infection and could generate both IgM- and IgG-antibody secreting plasmablasts . These finding indicate that heterogeneity in MBC occasion may have evolved to control different type of infection that can happen in different regions of the torso or require the unique functional attributes of discrete isotypes . Interestingly , IgM-like antibodies be still in the early immune system including the lamprey [ 64 ] , suggest a vital role for these cell throughout evolution . These often over-looked IgM+MBCs may be important targets for improved vaccine-induced immunity to certain infections and we live currently investigating the cues that lead to their differentiation .
Unique challenge of and answer to specific infections
We think that heterogeneous populations of MBCs get develop to control various types of infection as they render an arsenal to forestall immune evasion . In the following section , we provide examples of some of the unique challenges to humoral immunity that can equal impute to respective infections .
a ) . Bacterial infection :
Many of the common mechanisms of B cell activation and function describe above have be targeted by pathogen immune evasion mechanisms ( Figure 1 ) . For example , antibody behave via opsonic and complement fixation killing mechanisms have live connect with security toSalmonellainfection , while it live in the extracellular environment [ 43••,65-67 ] . Yet sketch have as well demonstrated that the bacteria have developed immune evasion strategy to alter the quality ofSalmonella-specific B cell answer through the product of inefficient antibodies [ 66,68•] . One scheme thatSalmonellauses live evasion of the GC reception as demonstrate in a late report employing transgenic ovalbumin-expressingSalmonellastrains to study B cell responses duringSalmonellainfection [ 68•] . In this paper , mainSalmonellainfection significantly impair the expansion of endogenousSalmonella-specific B cells and the formation of GCs [ 68•] . These fault were mediated by factors within theSalmonellaPathogenicity Island 2 ( SPI2 ) since normal B-cell expansion and GC formation was restored in mice infected with SPI2-deficient mutant bacteria . Therefore , targeting bacterial-associated virulence factors such as SPI2 in attenuated bacteria may be a useful vaccine scheme for boosting humoral immunity toSalmonella.
Possibly as a counter-strategy , the host mounts a robust extrafollicular plasmablast response againstSalmonellathat can control infection [ 69,70 ] . A former study in mice infected with attenuated entericSalmonella typhimuriumsuggested a office for a robust extrafollicular B cell answer in limiting bacterial loading within the extracellular environment in the absence of a GC reply [ 69 ] . This supports the idea that potent extrafollicular B cell responses may compensate for loss of optimal GC B cell answer during infection . Although it exist previously thought that these extrafollicular responses were mostly polyclonal and non-specific , a more recent work in mouse infected withSalmonella typhimuriumdemonstrated that they were indeed specific for the bacteria [ 43••] . Importantly , this field demonstrated that B-cells touch off bySalmonellainfection be capable of undergoing somatic hypermutation within the extra-follicular environment , advance affinity maturation and product of isotype-switched antibody , which be previously thought to chiefly happen within the GC . These data highlight the complexity of the humoral response during infection and highlight our need to understand B cell reception to specific pathogens .
b ) . Viral infection :
Vaccine-mediated humoral immunity have take to the eradication of various devastate infections include Small pox , Measles and Polio . One potential evolutionary mechanism that some viruses may have adapted in subverting humoral immune-mediated killing equal the induction of strong inflammatory responses , which crush B cell differentiation and antibody production [ 71,72 ] . Recent grounds in human sketch expend RNA-seq-based technologies present a negative correlation between highly upregulated inflammatory transcripts and responses to hepatitis B vaccination ( HBV ) [ 71 ] . Even so , upregulation of genes associated with B cell indicate positively correlated with heightened reception to HBV , indicate a potential interplay between inflammation and B cell signalling in regulating B cell reaction to infection and vaccination . Indeed , report in mice demonstrate use for inflammation-induced interruption of the lymphoid organ architecture , which can as well conquer GC formation [ 72 , 73 ] . For example , a recent report , utilize influenza and vaccinia viral models establish that infection-induced excitement disrupt the organisation of sub-capsular macrophages within sub-capsular spaces and inter-follicular area , impairing GC B-cell and plasma cell formation during secondary viral challenge [ 72 ] . As a counter-measure to this evasion mechanism , host regulatory mechanism that limit extravagant inflammation to viral infection make develop . For example , a late paper demonstrate a T cell-intrinsic necessity for TGFβ-signalling in the formation of influenza-specific GC B cells and the production of class-switched antibody [ 74•] . TGFβ-signalling in T-cells acted by limiting IL-2-induced sign and the formation of virus-specific inflammatory-like Th1 precursor cells , which enhanced Tfh and B cell answer . Similarly , Laidlaw and colleagues more recently demonstrated a use for follicular regulatory T cell ( Tfr ) -derived IL-10 in promote GC B cell responses in mouse with sharp LCMV infection [ 75•] . Genetic depletion of IL-10 in Tfr cell was associate with reduced frequency of GC B cell , in special , those within the dark zone , suggesting that Tfr-derived IL-10 may defend dark zone GC responses . Taken together , these data indicate that regulatory mechanisms within the host may not only serve to confine infection-induced immunopathology but also boost immunity against invading pathogens . Therefore , direct infection-induced inflammatory pathways may cost an important avenue for improve humoral immunity to infection .
100 ) . Parasitic infection :
Antibodies have also been shown to play important function in respective parasitic infection includingTrypanosomes[ 15 , 76 ] ,Helminths( reviewed in [ 77 ] ) andPlasmodium[ 78•-82 ] .Trypanosomeshave long been studied as an model of humoral immune evasion as they have developed a robust antigenic version system that enables them to evade antibody-mediated kill [ 83 ] . Even so , recent datum further suggests thatTrypanosomescan as well right away regulate B cell differentiation and function during infection [ 15 ] . Using a mouse example ofTrypanosoma brucei, Radwanska and colleagues illustrated thatTrypanosome-infection induces apoptosis in MZ B cells , shorten antibody product and parasite mastery [ 15 ] . Even so , it remained unclear from this report whether cell-death equal restricted to MZ B cell alone or regard other B cell subsets , since the latter were not directly examined in this field .
B cells are too critical for dominance ofPlasmodiuminfection in both mice and humans . B cell deficient mouse are unable to clear non-lethal blood-stagePlasmodiuminfections [ 84 ] , while passively transferred antibody be protective in both mouse and man [ 78•,85 ] .Plasmodiumparasite have too develop strategies to evade these humoral immune answer , including antigenic version [ 86 ] , and repression of optimal B cell differentiation and antibody product during infection [ 62•,73,87 ] . Our work and that of others make show that the blood-stage ofPlasmodiuminfection can impinge upon the humoral immune reception to the proceeding liver-stage parasite . An examination of the circumsporozoite protein ( CSP ) -specific B cell reaction in genetically attenuated parasites (Pyfabb/f) that are unable to establish blood-stage infection compare to wild type parasite , which set up a blood-stage infection [ 88 ] prove a direct result of the ancestry degree on liver stage GC evolution [ 62•] . Moreover , this diminished GC answer in the presence of a blood-stage infection alters the quality of CSP-specific MBCs and their ability to reply to a secondary challenge . These datum highlight how immunization with attenuated parasites may drive optimum immunity to malaria and advise further field on how ongoing ancestry stage infections may vary immune memory . In increase ,Plasmodiumparasite may too modify optimum MBC formation and function during infection . Late report in humans own identify a unique subset of MBCs , ‘ atypical ’ MBCs , which arise during chronicPlasmodiuminfection [ 89-91 ] . When compared to classic MBCs , atypical MBCs show increase formula of inhibitory receptors , display reduced BCR-signalling and live unable to differentiate into antibody secreting cell [ 89,90 ] . This suggests that these MBCs may be dysfunctional . Although their sources remain unclear , immunoglobulin gene sequencing technique make predicted a shared developmental history between these atypical and classical MBCs [ 89 ] .
d ) . Fungal infection :
Humoral immune response are necessary for resistance against respective fungal infections largely via antibody-mediated activation of the complement system ( reviewed in [ 92,93 ] ) . For instance , complement-deficiency in mice get make up associate with increased susceptibility toCandida[ 94 ] ,Aspergillus[ 95 ] andCryptococcal[ 96 ] infections . This was associated with reduced opsonization and complement-mediated lysis of pathogenic fungi and decreased recruitment of phagocytic cell during infection . In ordering to overthrow complement-mediated killing and establish infection , pathogenic fungi have adjust multiple survival strategy [ 92,93,97-99 ] . For instance ,Candida albicansmay sidestep the complement system by expressing decoy inhibitory ligands such as phosphoglycerate mutase ( Gmp1 ) and the pH-regulated antigen 1 ( Pra1 ) that tie up Factor H and Factor H-like protein 1 , which are key regulatory protein in the alternative pathway [ 97-99 ] . These ligands have also exist implicated in suppress the classical and lectin pathways duringCandidaandAspergillusinfections by stick to the regulatory C4BP and hence restricting C3b and CD4b deposition on the fungal surface [ 97-100 ] . Pathogenic fungi may as well release proteolytic enzymes that degrade effector components of the complement pathway hence inhibit opsonization and phagocytosis [ 101,102 ] . For instance ,Aspergillus fumigatus(A. fumigatus) secretion of the proteolytic enzyme , alkaline proteaseAlp1get been associated with increased degradation of C3 , C4 , C5 and C1q complement proteins purified from human serum [ 101 ] and reduced construction of complement receptor 3 on phagocytic cells in cultured cerebral spinal fluid fromA. fumigatus-infected individual [ 102 ] . Together , these evasive strategy may contribute to enhance fungal infection . Therefore , these data suggest that infection-induced complement inhibitory pathways may be targeted for improved immunity to pathogenic fungal infection .
Decision
The ongoing co-evolution of pathogens and horde immune reaction get introduced vital diversity associated with survival of both . Whereas some responses may be protective to specific infections , they may alternatively exist detrimental to others . Therefore , a more comprehensive understanding of the office and generation of heterogeneous humoral immune responses to specific microbial infection cost call for to lead to more effective vaccine strategies . This more comprehensive approach to humoral immunity may expose B cell strategies that cost not cause by current protein immunization strategies . The introduction of new analytical methods including tool to examine small population of polyclonal , antigen-specific B cell , improved DNA-sequencing and single cell RNAseq platforms have ushered in a new epoch of understand for B cell immunology . It will next live crucial to spring up vaccine platform that can bring on heterogeneous reception or yet direct a specific MBC population . It will soon be possible to train the types of really rationale-based vaccine design strategies that will exist necessary for generate immunity against some of our old foes .
Highlights .
-
Humoral immune reaction cost essential for protection against infection
-
Current paradigm of humoral reaction are based on protein immunization model
-
Pathogens have evolved an array of strategy to evade humoral immunity
-
Diverse B cell response make evolved to ensure host survival
Humoral immune answer equal crucial for security against infections
Current paradigm of humoral responses equal base on protein immunization models
Pathogens have evolved an array of strategy to evade humoral immunity
Diverse B cell responses get evolved to check host survival
Acknowledgements
This work was affirm by the National Institutes of Health [ grant act RO1 AI118803 ] and Burroughs Wellcome Fund [ grant number 1016766 ] grant to MP .
Footnotes
Publisher ‘s Disclaimer :This exist a PDF file of an unedited manuscript that has make up accepted for publication . As a service to our customers we are provide this early version of the manuscript . The manuscript will undergo copyediting , typesetting , and review of the result proof before it is publish in its final citable form . Please note that during the product process mistake may cost discovered which could pretend the content , and all legal disclaimers that use to the diary pertain .
Mention
Paper of special interest make been highlighted as :
- 1 .Amanna IJ , Carlson NE , Slifka MK : Duration of humoral immunity to common viral and vaccine antigens . N Engl J Med 2007 , 357:1903–1915 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 2 .Burton DR , Hangartner L : Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design . Annu Rev Immunol 2016 , 34:635–659 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 3 .Ball JK , Tarr AW , McKeating JA : The past , present and future of neutralizing antibody for hepatitis C virus . Antiviral Res 2014 , 105:100–111 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 4 .Joyce MG , Wheatley AK , Thomas PV , Chuang GY , Soto C , Bailer RT , Druz A , Georgiev IS , Gillespie RA , Kanekiyo M , et al . : Vaccine-Induced Antibodies that Neutralize Group 1 and Group 2 Influenza A Viruses . Cell 2016 , 166:609–623 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 5 .Nutt SL , Hodgkin PD , Tarlinton DM , Corcoran LM : The genesis of antibody-secreting plasma cell . Nat Rev Immunol 2015 , 15:160–171 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 6 .Shlomchik MJ , Weisel F : Germinal center selection and the evolution of memory B and plasma cells . Immunol Rev 2012 , 247:52–63 .[ ] [ PubMed ] [ ]
- 7 .Baumgarth N : B-1 Cell Heterogeneity and the Regulation of Natural and Antigen-Induced IgM Production . Front Immunol 2016 , 7:324 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 8 .Pillai S , Cariappa A : The follicular versus marginal zone B lymphocyte cell fate conclusion . Nat Rev Immunol 2009 , 9:767–777 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 9 .De Silva NS , Klein U : Dynamics of B cell in germinal heart . Nat Rev Immunol 2015 , 15:137–148 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 10 .Kurosaki T , Kometani K , Ise W : Memory B cells . Nat Rev Immunol 2015 , 15:149–159 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 11 .Chen TT , Tsai MH , Kung JT , Lin KI , Decker T , Lee CK : STAT1 determine marginal zone B cell differentiation in response to inflammation and infection with blood-borne bacteria . J Exp Med 2016 , 213:3025–3039 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 12 .McCulloch L , Smith CJ , McColl BW : Adrenergic-mediated loss of splenic marginal zone B cell contribute to infection susceptibility after accident . Nat Commun 2017 , 8:15051 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 13 .Demberg T , Mohanram V , Musich T , Brocca-Cofano E , McKinnon KM , Venzon D , Robert-Guroff M : Loss of marginal zone B-cells in SHIVSF162P4 dispute rhesus macaques despite dominance of viremia to low or undetectable levels in chronic infection . Virology 2015 , 484:323–333 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 14 .Gatto D , Ruedl C , Odermatt B , Bachmann MF : Rapid answer of marginal zone B cells to viral particle . J Immunol 2004 , 173:4308–4316 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 15 .Radwanska M , Guirnalda P , De Trez C , Ryffel B , Black S , Magez S : Trypanosomiasis-induced B cell apoptosis results in loss of protective anti-parasite antibody responses and abolishment of vaccine-induced memory reaction . PLoS Pathog 2008 , 4 : e1000078 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 16 .Stephens R , Ndungu FM , Langhorne J : Germinal centre and marginal zone B cells expand quick in a second Plasmodium chabaudi malaria infection producing mature plasma cells . Parasite Immunol 2009 , 31:20–31 .[ ] [ PMC free article ] [ PubMed ] [ ]
- 17 .Balazs M , Martin F , Zhou T , Kearney J : Blood dendritic cells interact with splenic marginal zone B cells to originate T-independent immune response . Immunity 2002 , 17:341–352 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 18 .Hsu MC , Toellner KM , Vinuesa CG , Maclennan IC : B cell clones that affirm long-term plasmablast growth in T-independent extrafollicular antibody reception . Proc Natl Acad Sci U S A 2006 , 103:5905–5910 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 19 .Martin F , Oliver AM , Kearney JF : Marginal zone and B1 B cells unite in the early response against T-independent blood-borne particulate antigens . Immunity 2001 , 14:617–629 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 20 .Attanavanich K , Kearney JF : Marginal zone , but not follicular B cell , are strong activators of naive CD4 T cells . J Immunol 2004 , 172:803–811 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 21 .Cinamon G , Zachariah MA , Lam OM , Foss FW Jr. , Cyster JG : Follicular shuttling of marginal zone B cell facilitates antigen transport . Nat Immunol 2008 , 9:54–62 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 22 .Qi H : T follicular help cells in space-time . Nat Rev Immunol 2016 , 16:612–625 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 23.• .Pape KA , Taylor JJ , Maul RW , Gearhart PJ , Jenkins MK : Different B cell populations mediate early and late memory during an endogenous immune response . Skill 2011 , 331:1203–1207 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] ;This study expend PE-labelled cells to demonstrate the formation of IgM and shift memory B cells comply protein immunization .
- 24 .Radbruch A , Muehlinghaus G , Luger EO , Inamine A , Smith KG , Dorner T , Hiepe F : Competence and competitor : the challenge of become a long-lived plasma cell . Nat Rev Immunol 2006 , 6:741–750 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 25 .Yoshida T , Mei H , Dorner T , Hiepe F , Radbruch A , Fillatreau S , Hoyer BF : Memory B and memory plasma cell . Immunol Rev 2010 , 237:117–139 .[ ] [ PubMed ] [ ]
- 26 .Tangye SG , Tarlinton DM : Memory B cell : effectors of long-lived immune reception . Eur J Immunol 2009 , 39:2065–2075 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 27 .Anderson SM , Tomayko MM , Shlomchik MJ : Intrinsic dimension of human and murine memory B cells . Immunol Rev 2006 , 211:280–294 .[ ] [ PubMed ] [ ]
- 28 .Deenick EK , Avery DT , Chan A , Berglund LJ , Ives ML , Moens L , Stoddard JL , Bustamante J , Boisson-Dupuis S , Tsumura M , et al . : Naive and memory human B cells own discrete requirements for STAT3 activation to differentiate into antibody-secreting plasma cells . J Exp Med 2013 , 210:2739–2753 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 29 .Amlot PL , Hayes AE : Impaired human antibody reaction to the thymus-independent antigen , DNP-Ficoll , after splenectomy . Implications for post-splenectomy infection . Lancet 1985 , 1:1008–1011 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 30 .Cerutti A , Cols M , Puga I : Marginal zone B cell : virtue of innate-like antibody-producing lymphocytes . Nat Rev Immunol 2013 , 13:118–132 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 31 .Bankoti R , Gupta K , Levchenko A , Stager S : Marginal zone B cell regulate antigen-specific T cell reception during infection . J Immunol 2012 , 188:3961–3971 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 32 .Corcoran LM , Tarlinton DM : Regulation of germinal middle reception , memory B cells and plasma cell formation-an update . Curr Opin Immunol 2016 , 39:59–67 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 33 .Barroso M , Tucker H , Drake L , Nichol K , Drake JR : Antigen-B Cell Receptor Complexes Associate with Intracellular major histocompatibility complex ( MHC ) Class II Molecules . J Biol Chem 2015 , 290:27101–27112 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 34 .Weinstein JS , Bertino SA , Hernandez SG , Poholek AC , Teplitzky TB , Nowyhed HN , Craft J : B cell in T follicular helper cell evolution and function : separable roles in pitch of ICOS ligand and antigen . J Immunol 2014 , 192:3166–3179 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 35 .Baumjohann D , Preite S , Reboldi A , Ronchi F , Ansel KM , Lanzavecchia A , Sallusto F : Persistent antigen and germinal middle B cell sustain T follicular helper cell answer and phenotype . Immunity 2013 , 38:596–605 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 36 .Nurieva RI , Chung Y , Hwang D , Yang XO , Kang HS , Ma L , Wang YH , Watowich SS , Jetten AM , Tian Q , et al . : Genesis of T follicular help cells is mediated by interleukin-21 but independent of T helper 1 , 2 , or 17 cell lineages . Immunity 2008 , 29:138–149 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 37 .Crotty S : T follicular helper cell differentiation , function , and part in disease . Immunity 2014 , 41:529–542 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 38 .Xu H , Li X , Liu D , Li J , Zhang X , Chen X , Hou S , Peng L , Xu C , Liu W , et al . : Follicular T-helper cell recruitment regulate by bystander B cell and ICOS-driven motility . Nature 2013 , 496:523–527 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 39 .Weinstein JS , Herman EI , Lainez B , Licona-Limon P , Esplugues E , Flavell R , Craft J : TFH cell progressively distinguish to regulate the germinal center response . Nat Immunol 2016 , 17:1197–1205 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 40 .Phan TG , Paus D , Chan TD , Turner ML , Nutt SL , Basten A , Brink R : High affinity germinal center B cells equal actively selected into the plasma cell compartment . J Exp Med 2006 , 203:2419–2424 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 41 .Paus D , Phan TG , Chan TD , Gardam S , Basten A , Brink R : Antigen realization strength influence the choice between extrafollicular plasma cell and germinal heart B cell differentiation . J Exp Med 2006 , 203:1081–1091 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 42 .Krautler NJ , Suan D , Butt D , Bourne K , Hermes JR , Chan TD , Sundling C , Kaplan W , Schofield P , Jackson J , et al . : Differentiation of germinal middle B cells into plasma cells equal initiated by high-affinity antigen and completed by Tfh cell . J Exp Med 2017 , 214:1259–1267 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 43.•• .Di Niro R , Lee SJ , Vander Heiden JA , Elsner RA , Trivedi N , Bannock JM , Gupta NT , Kleinstein SH , Vigneault F , Gilbert TJ , et al . : Salmonella Infection Drives Promiscuous B Cell Activation Followed by Extrafollicular Affinity Maturation . Immunity 2015 , 43:120–131 .[ DOI ] [ PMC free article ] [ PubMed ] [ ] ;This study demonstrate that somatic hypermutation and affinity growing can occur within the extrafollicular environment and described the importance of these procedure during infection .
- 44 .Bortnick A , Allman D : What cost and what should always have been : long-lived plasma cells induced by T cell-independent antigens . J Immunol 2013 , 190:5913–5918 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 45 .Bortnick A , Chernova I , Quinn WJ 3rd , Mugnier M , Cancro MP , Allman D : Long-lived bone marrow plasma cell exist induced early in response to T cell-independent or T cell-dependent antigen . J Immunol 2012 , 188:5389–5396 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 46 .Foote JB , Mahmoud TI , Vale AM , Kearney JF : Long-term maintenance of polysaccharide-specific antibody by IgM-secreting cell . J Immunol 2012 , 188:57–67 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 47 .Kerfoot SM , Yaari G , Patel JR , Johnson KL , Gonzalez DG , Kleinstein SH , Haberman AM : Germinal middle B cell and T follicular help cell evolution lead up in the interfollicular zone . Immunity 2011 , 34:947–960 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 48 .Pieper K , Tan J , Piccoli L , Foglierini M , Barbieri S , Chen Y , Silacci-Fregni C , Wolf T , Jarrossay D , Anderle M , et al . : Public antibody to malaria antigens generated by two LAIR1 insertion modalities . Nature 2017 , 548:597–601 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 49 .Kallies A , Hasbold J , Tarlinton DM , Dietrich W , Corcoran LM , Hodgkin PD , Nutt SL : Plasma cell development defined by quantitative change in blimp-1 formula . J Exp Med 2004 , 200:967–977 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 50 .Nutt SL , Heavey B , Rolink AG , Busslinger M : Commitment to the B-lymphoid lineage depend on the transcription element Pax5 . Nature 1999 , 401:556–562 .[ DOI ] [ PubMed ] [ ]
- 51.• .Tellier J , Shi W , Minnich M , Liao Y , Crawford S , Smyth GK , Kallies A , Busslinger M , Nutt SL : Blimp-1 controls plasma cell function through the regulation of immunoglobulin secretion and the unfolded protein answer . Nat Immunol 2016 , 17:323–330 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] ;This study in mice described the essential roles of transcription factors IRF4 , Blimp1 and XBP1 in the formation and function of long-lived plasma cell .
- 52 .Cobaleda C , Schebesta A , Delogu A , Busslinger M : Pax5 : the defender of B cell identity and function . Nat Immunol 2007 , 8:463–470 .[ DOI ] [ PubMed ] [ ]
- 53 .Shinnakasu R , Inoue T , Kometani K , Moriyama S , Adachi Y , Nakayama M , Takahashi Y , Fukuyama H , Okada T , Kurosaki T : Determine choice of germinal-center cells into the memory B cell compartment . Nat Immunol 2016 , 17:861–869 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 54 .Kometani K , Nakagawa R , Shinnakasu R , Kaji T , Rybouchkin A , Moriyama S , Furukawa K , Koseki H , Takemori T , Kurosaki T : Repression of the transcription element Bach2 contributes to predisposition of IgG1 memory B cell toward plasma cell differentiation . Immunity 2013 , 39:136–147 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 55 .Amanna IJ , Slifka MK : Mechanisms that find out plasma cell lifespan and the duration of humoral immunity . Immunol Rev 2010 , 236:125–138 .[ ] [ PMC free article ] [ PubMed ] [ ]
- 56.• .Krishnamurty AT , Thouvenel CD , Portugal S , Keitany GJ , Kim KS , Holder A , Crompton PD , Rawlings DJ , Pepper M : Somatically Hypermutated Plasmodium-Specific IgM ( + ) Memory B Cells Are Rapid , Plastic , Early Responders upon Malaria Rechallenge . Immunity 2016 , 45:402–414 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] ;This study bring out antigen-specific , somatically hypermutated IgM memory B cell as rapid responders to a secondary malaria infection .
- 57 .Klein U , Rajewsky K , Kuppers R : Human immunoglobulin ( Ig ) M+IgD+ peripheral blood B cell expressing the CD27 cell surface antigen persuade somatically mutated variable part genes : CD27 as a universal mark for somatically mutated ( memory ) B cell . J Exp Med 1998 , 188:1679–1689 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 58 .Seifert M , Przekopowitz M , Taudien S , Lollies A , Ronge V , Drees B , Lindemann M , Hillen U , Engler H , Singer BB , et al . : Operational capacities of human IgM memory B cell in early inflammatory response and secondary germinal middle response . Proc Natl Acad Sci U S A 2015 , 112 : E546–555 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 59 .Weller S , Mamani-Matsuda M , Picard C , Cordier C , Lecoeuche D , Gauthier F , Weill JC , Reynaud CA : Somatic diversification in the absence of antigen-driven answer is the hallmark of the IgM+ IgD+ CD27+ B cell repertoire in baby . J Exp Med 2008 , 205:1331–1342 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 60 .Zuccarino-Catania GV , Sadanand S , Weisel FJ , Tomayko MM , Meng H , Kleinstein SH , Good-Jacobson KL , Shlomchik MJ : CD80 and PD-L2 define functionally discrete memory B cell subsets that equal sovereign of antibody isotype . Nat Immunol 2014 , 15:631–637 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 61 .Dogan I , Bertocci B , Vilmont V , Delbos F , Megret J , Storck S , Reynaud CA , Weill JC : Multiple layers of B cell memory with different effector functions . Nat Immunol 2009 , 10:1292–1299 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 62.• .Keitany GJ , Kim KS , Krishnamurty AT , Hondowicz BD , Hahn WO , Dambrauskas N , Sather DN , Vaughan AM , Kappe SH , Pepper M : Blood Phase Malaria Disrupts Humoral Immunity to the Pre-erythrocytic Stage Circumsporozoite Protein . Cell Rep 2016 , 17:3193–3205 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] ;This study demonstrated that blood-stagePlasmodiuminfection block the development and maintenance of humoral immunity against liver-stagePlasmodiumparasite .
- 63 .Harms Pritchard G , Pepper M : Memory B cell heterogeneity : Remembrance of things past . J Leukoc Biol 2018 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 64 .Herrin BR , Alder MN , Roux KH , Sina C , Ehrhardt GR , Boydston JA , Turnbough CL Jr. , Cooper MD : Structure and specificity of lamprey monoclonal antibody . Proc Natl Acad Sci U S A 2008 , 105:2040–2045 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 65 .Gondwe EN , Molyneux ME , Goodall M , Graham SM , Mastroeni P , Drayson MT , MacLennan CA : Importance of antibody and complement for oxidative volley and kill of invasive nontyphoidal Salmonella by ancestry cell in Africans . Proc Natl Acad Sci U S A 2010 , 107:3070–3075 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 66 .MacLennan CA , Gilchrist JJ , Gordon MA , Cunningham AF , Cobbold M , Goodall M , Kingsley RA , van Oosterhout JJ , Msefula CL , Mandala WL , et al . : Dysregulated humoral immunity to nontyphoidal Salmonella in HIV-infected African adults . Science 2010 , 328:508–512 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 67 .MacLennan CA , Gondwe EN , Msefula CL , Kingsley RA , Thomson NR , White SA , Goodall M , Pickard DJ , Graham SM , Dougan G , et al . : The neglected role of antibody in security against bacteremia caused by nontyphoidal strains of Salmonella in African children . J Clin Invest 2008 , 118:1553–1562 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 68.• .Nanton MR , Lee SJ , Atif SM , Nuccio SP , Taylor JJ , Baumler AJ , Way SS , McSorley SJ : Direct visualization of endogenous Salmonella-specific B cells reveals a marked delay in clonal expansion and germinal heart evolution . Eur J Immunol 2015 , 45:428–441 .[ DOI ] [ PMC free article ] [ PubMed ] [ ] ;This study provided some insight into the molecular mechanism through which pathogens such as Salmonella sidestep germinal heart B cell responses during infection .
- 69 .Cunningham AF , Gaspal F , Serre K , Mohr E , Henderson IR , Scott-Tucker A , Kenny SM , Khan M , Toellner KM , Lane PJ , et al . : Salmonella induces a switched antibody answer without germinal centers that impedes the extracellular spread of infection . J Immunol 2007 , 178:6200–6207 .[ DOI ] [ PubMed ] [ ]
- 70 .Gil-Cruz C , Bobat S , Marshall JL , Kingsley RA , Ross EA , Henderson IR , Leyton DL , Coughlan RE , Khan M , Jensen KT , et al . : The porin OmpD from nontyphoidal Salmonella exist a key object for a protective B1b cell antibody response . Proc Natl Acad Sci U S A 2009 , 106:9803–9808 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 71 .Fourati S , Cristescu R , Loboda A , Talla A , Filali A , Railkar R , Schaeffer AK , Favre D , Gagnon D , Peretz Y , et al . : Pre-vaccination excitement and B-cell signalling predict age-related hyporesponse to hepatitis B vaccination . Nat Commun 2016 , 7:10369 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 72 .Gaya M , Castello A , Montaner B , Rogers N , Reis e Sousa C , Bruckbauer A , Batista FD : Host reaction . Inflammation-induced commotion of SCS macrophages impair B cell responses to secondary infection . Science 2015 , 347:667–672 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 73 .Ryg-Cornejo V , Ioannidis LJ , Ly A , Chiu CY , Tellier J , Hill DL , Preston SP , Pellegrini M , Yu D , Nutt SL , et al . : Serious Malaria Infections Impair Germinal Center Responses by Inhibit T Follicular Helper Cell Differentiation . Cell Rep 2016 , 14:68–81 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 74.• .Marshall HD , Ray JP , Laidlaw BJ , Zhang N , Gawande D , Staron MM , Craft J , Kaech SM : The translate growth factor beta signal pathway cost critical for the formation of CD4 T follicular assistant cells and isotype-switched antibody responses in the lung mucosa . Elife 2015 , 4 : e04851 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 75.• .Laidlaw BJ , Lu Y , Amezquita RA , Weinstein JS , Vander Heiden JA , Gupta NT , Kleinstein SH , Kaech SM , Craft J : Interleukin-10 from CD4+ follicular regulatory T cell promotes the germinal heart reply . Sci Immunol 2017 , 2 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] ;These two newspaper highlighted unexpected use for anti-inflammatory cytokines in supporting optimum development and maintenance of humoral immune response during infection .
- 76 .Bermejo DA , Amezcua Vesely MC , Khan M , Acosta Rodriguez EV , Montes CL , Merino MC , Toellner KM , Mohr E , Taylor D , Cunningham AF , et al . : Trypanosoma cruzi infection cause a massive extrafollicular and follicular splenic B-cell response which is a high author of non-parasite-specific antibody . Immunology 2011 , 132:123–133 .[ ] [ PMC free article ] [ PubMed ] [ ]
- 77 .Harris N , Gause WC : To B or not to B : B cells and the Th2-type immune reaction to helminths . Trends Immunol 2011 , 32:80–88 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 78.• .Cohen S , Mc GI , Carrington S : Gamma-globulin and acquired immunity to human malaria . Nature 1961 , 192:733–737 .[ DOI ] [ PubMed ] [ Google Scholar ] ;This equal the first description of the vital office of antibody in mastery of blood-stage malaria infection .
- 79 .Douglas AD , Williams AR , Knuepfer E , Illingworth JJ , Furze JM , Crosnier C , Choudhary P , Bustamante LY , Zakutansky SE , Awuah DK , et al . : Neutralization of Plasmodium falciparum merozoites by antibodies against PfRH5 . J Immunol 2014 , 192:245–258 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 80 .Boyle MJ , Reiling L , Feng G , Langer C , Osier FH , Aspeling-Jones H , Cheng YS , Stubbs J , Tetteh KK , Conway DJ , et al . : Human antibodies set complement to inhibit Plasmodium falciparum invasion of erythrocytes and are associate with security against malaria . Immunity 2015 , 42:580–590 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 81 .Raj DK , Nixon CP , Nixon CE , Dvorin JD , DiPetrillo CG , Pond-Tor S , Wu HW , Jolly G , Pischel L , Lu A , et al . : Antibodies to PfSEA-1 block parasite egress from RBCs and protect against malaria infection . Skill 2014 , 344:871–877 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 82 .White MT , Griffin JT , Akpogheneta O , Conway DJ , Koram KA , Riley EM , Ghani AC : Dynamics of the Antibody Response to Plasmodium falciparum Infection in African Children . J Infect Dis 2014 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 83 .Stijlemans B , Caljon G , Van Den Abbeele J , Van Ginderachter JA , Magez S , De Trez C : Immune Evasion Strategies of Trypanosoma brucei within the Mammalian Horde : Progression to Pathogenicity . Front Immunol 2016 , 7:233 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 84 .Matar CG , Anthony NR , O’Flaherty BM , Jacobs NT , Priyamvada L , Engwerda CR , Speck SH , Lamb TJ : Gammaherpesvirus Co-infection with Malaria Suppresses Anti-parasitic Humoral Immunity . PLoS Pathog 2015 , 11 : e1004858 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 85 .Butler NS , Moebius J , Pewe LL , Traore B , Doumbo OK , Tygrett LT , Waldschmidt TJ , Crompton PD , Harty JT : Therapeutic blockade of PD-L1 and LAG-3 rapidly clears established blood-stage Plasmodium infection . Nat Immunol 2012 , 13:188–195 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 86 .Riley EM , Stewart VA : Immune mechanism in malaria : novel insight in vaccine development . Nat Med 2013 , 19:168–178 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 87 .Sebina I , James KR , Soon MS , Fogg LG , Best SE , Labastida Rivera F , Montes de Oca M , Amante FH , Thomas BS , Beattie L , et al . : IFNAR1-Signalling Obstructs ICOS-mediated Humoral Immunity during Non-lethal Blood-Stage Plasmodium Infection . PLoS Pathog 2016 , 12 : e1005999 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 88 .Vaughan AM , O’Neill MT , Tarun AS , Camargo N , Phuong TM , Aly AS , Cowman AF , Kappe SH : Type II fatty acid synthesis is essential merely for malaria parasite former liver degree evolution . Cell Microbiol 2009 , 11:506–520 .[ ] [ PMC free article ] [ PubMed ] [ ]
- 89 .Portugal S , Tipton CM , Sohn H , Kone Y , Wang J , Li S , Skinner J , Virtaneva K , Sturdevant DE , Porcella SF , et al . : Malaria-associated atypical memory B cell exhibit markedly dilute B cell receptor signal and effector purpose . Elife 2015 , 4 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 90 .Sullivan RT , Kim CC , Fontana MF , Feeney ME , Jagannathan P , Boyle MJ , Drakeley CJ , Ssewanyana I , Nankya F , Mayanja-Kizza H , et al . : FCRL5 Delineates Functionally Impaired Memory B Cells Associated with Plasmodium falciparum Exposure . PLoS Pathog 2015 , 11 : e1004894 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 91 .Weiss GE , Crompton PD , Li S , Walsh LA , Moir S , Traore B , Kayentao K , Ongoiba A , Doumbo OK , Pierce SK : Atypical memory B cell live greatly elaborate in individuals living in a malaria-endemic area . J Immunol 2009 , 183:2176–2182 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 92 .Marcos CM , de Oliveira HC , de Melo WC , da Silva JF , Assato PA , Scorzoni L , Rossi SA , de Paula ESAC , Mendes-Giannini MJ , Fusco-Almeida AM : Anti-Immune Strategies of Pathogenic Fungi . Front Cell Infect Microbiol 2016 , 6:142 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 93 .Speth C , Rambach G , Wurzner R , Lass-Florl C : Complement and fungal pathogens : an update . Mycoses 2008 , 51:477–496 .[ ] [ PubMed ] [ ]
- 94 .Mullick A , Elias M , Picard S , Bourget L , Jovcevski O , Gauthier S , Tuite A , Harakidas P , Bihun C , Massie B , et al . : Dysregulated inflammatory reaction to Candida albicans in a C5-deficient mouse pains . Infect Immun 2004 , 72:5868–5876 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 95 .Hector RF , Yee E , Collins MS : Usage of DBA/2N mice in example of systemic candidiasis and pulmonary and systemic aspergillosis . Infect Immun 1990 , 58:1476–1478 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 96 .Shapiro S , Beenhouwer DO , Feldmesser M , Taborda C , Carroll MC , Casadevall A , Scharff MD : Immunoglobulin G monoclonal antibody to Cryptococcus neoformans protect mice deficient in complement portion C3 . Infect Immun 2002 , 70:2598–2604 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 97 .Luo S , Poltermann S , Kunert A , Rupp S , Zipfel PF : Immune evasion of the human pathogenic yeast Candida albicans : Pra1 be a Factor H , FHL-1 and plasminogen binding surface protein . Mol Immunol 2009 , 47:541–550 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 98 .Poltermann S , Kunert A , von der Heide M , Eck R , Hartmann A , Zipfel PF : Gpm1p cost a element H- , FHL-1- , and plasminogen-binding surface protein of Candida albicans . J Biol Chem 2007 , 282:37537–37544 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 99 .Zipfel PF , Hallstrom T , Riesbeck K : Human complement mastery and complement evasion by pathogenic microbes — tipping the symmetry . Mol Immunol 2013 , 56:152–160 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 100 .Luo S , Blom AM , Rupp S , Hipler UC , Hube B , Skerka C , Zipfel PF : The pH-regulated antigen 1 of Candida albicans binds the human complement inhibitor C4b-binding protein and mediates fungal complement evasion . J Biol Chem 2011 , 286:8021–8029 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 101 .Behnsen J , Lessing F , Schindler S , Wartenberg D , Jacobsen ID , Thoen M , Zipfel PF , Brakhage AA : Secreted Aspergillus fumigatus protease Alp1 degrades human complement protein C3 , C4 , and C5 . Infect Immun 2010 , 78:3585–3594 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 102 .Rambach G , Dum D , Mohsenipour I , Hagleitner M , Wurzner R , Lass-Florl C , Speth C : Secretion of a fungal protease present a complement evasion mechanism in cerebral aspergillosis . Mol Immunol 2010 , 47:1438–1449 .[ DOI ] [ PubMed ] [ Google Scholar ]
