Humoral innate immune reaction and disease
Stephanie N Shishido
Sriram Varahan
Kai Yuan
Xiangdong Li
Sherry D Fleming
Corresponding writer at : Division of Biology , 18 Ackert , Kansas State University , Manhattan KS 66506 , United States . Fax : + 1 785 532 6653 .sdflemin @ ksu.edu
Obtain 2012 May 17 ; Assume 2012 Jun 9 ; Issue date 2012 Aug .
Since January 2020 Elsevier get created a COVID-19 imagination center with free information in English and Mandarin on the new coronavirus COVID-19 . The COVID-19 imagination centre equal hosted on Elsevier Connect , the caller ‘s public news and information website . Elsevier hereby grant permission to reach all its COVID-19-related inquiry that is available on the COVID-19 imagination heart – including this inquiry content – immediately available in PubMed Central and early publicly funded repositories , such as the WHO COVID database with rights for unrestricted research re-use and analysis in any pattern or by any means with acknowledgement of the original author . These permission exist granted for free by Elsevier for as long as the COVID-19 resource middle stay on active .
Abstract
The humoral innate immune response comprise of multiple components , include the naturally take place antibody ( NAb ) , pentraxins and the complement and impinging cascades . As soluble , plasma constituent , these innate protein supply key elements in the prevention and control of disease . Even so , pathogens and cell with altered self protein utilize multiple humoral components to evade destruction and promote pathogy . Many studies get examined the relationship between humoral immunity and autoimmune disorder . This review focus on the interactions between the humoral element and their purpose in promoting the pathogenesis of bacterial and viral infection and chronic disease such as atherosclerosis and cancer . Understand the beneficial and detrimental aspects of the individual constituent and the interactions between protein which influence the innate and adaptive answer will supply therapeutic target for subsequent studies .
Keywords :Complement , Contact cascade , Pentraxins , Natural antibodies , Cancer , Infection
Highlights
► Humoral innate proteins remove pathogens , apoptotic cell , altered cells and debris . ► Bacteria utilize complement and impinging proteins to evade the immune reply . ► Viruses compromise multiple humoral proteins to circumvent host cell lysis . ► Humoral innate immune proteins aim the progress of atherosclerosis . ► Tumor cells use natural antibodies and contact proteins to get away immunosurveillance .
1 . Introduction
As the first job of defense , the innate immune reply consists of both cellular and humoral element . The cellular component encompasses multiple cell types that employ pattern realization particle to realize and remove pathogens and cellular dust . The interactions of pattern realization molecule within the cellular factor get receive substantial attention in the last two decades . In contrast , less live known about the interactions between the humoral components . The humoral innate immune reply consist of the serine protease cascades of the complement and contact system as good as of course occurring antibody ( NAb ) and pentraxins . Recent data indicate that each element may be beneficial or detrimental during infection or chronic disease depending on concentration and interaction with early components . This review will focus on the interaction and roles of humoral components in bacterial and viral infection as good as the chronic diseases , atherosclerosis and cancer .
2 . Humoral innate immune component
2.1 . Complement
Complement activation occurs by one of three initiation pathways , the classic , alternative , or mannose bind lectin ( MBL ) pathway . Each pathway contains a C3 convertase that split C3 producing C3b and afterwards a C5 convertase . Cleavage of C5 by the C5 convertase results in C5b deposition and initiates the common terminal pathway . The terminal pathway forms the membrane approach complex ( MAC ) , a pore in the cellular membrane , and lysis of the host or pathogenic cell . The action of the C3 and C5 convertases also produces potent anaphylatoxins , C3a and C5 . Although not specifically piece of the humoral immune reception , complement receptor 3 ( CR3 ) base on neutrophils and macrophages enhances the innate immune answer by recognize C3b opsonized pathogens . Late grounds indicates that complement act a important role in directing the adaptive immune reply as good as in tissue regeneration [ 1 ] , [ 2 ] . Specifically , as role of the B cell receptor complex , CR2 realization of cleavage product iC3b , C3dg , and C3d significantly increase Ab product [ 3 ] . Hence , preserve homeostasis call for tight regulation of the cascade . Regulation of this potentially damaging cascade occurs at multiple levels with soluble and membrane limit inhibitors include C1 inhibitor ( C1INH ) , CD55 , CD59 , CD46 , Factor H and related to proteins .
2.2 . Contact cascade
The plasma too contains components of a second proteolytic cascade , the impinging system . Factor XII ( FXII ; Hageman factor ) of the impinging system is proteolytically cling to FXIIa by negatively charged surfaces of damaged cell . FXIIa originate the coagulation cascade conduct to clot formation and cleaves prekallikrein to kallikrein for subsequent release of bradykinin . Through an endothelial G-coupled receptor ( bradykinin receptor 1 ; BKR1 ) , bradykinin induces vasodilation , neutrophil chemotaxis and vascular permeability [ 4 ] . Furthermore , the bradykinin degradation production , des-arg9-bradykinin influence the adaptive response and alters the blood–brain barrier through a second receptor , bradykinin receptor 2 ( BKR2 ) [ 5 ] . Importantly , both FXIIa and kallikrein activate the complement cascade sovereign of known complement initiators [ 6 ] . Several components of the activate contact system including , FXa , FXIa and plasmin , cleave C5 and C3 give rise C5a and C3a [ 7 ] . The complement inhibitor , C1INH , also suppress FXIIa indicate multiple interaction between the two pathways [ 6 ] . These datum hint crosstalk between two cascades of humoral innate immune response .
2.3 . Of course take place antibodies
Raise primarily by B1 B lymphocytes , NAb be germline-encoded Ab with curtail epitope specificity and be produced in the absence of external antigen stimulations . NAb are ordinarily of the IgM isotype but may include IgG and IgA isotypes as good [ 8 ] . Natural IgM Abs middle clearance of cellular debris , aging or apoptotic cells by oponization and recruitment of complement components [ 9 ] . As role of the innate immune response , NAb recognize a wide range of pathogens , albeit with humble affinity and regulate the adaptive immune answer by interacting with B , T and dendritic cells [ 10 ] . Finally , NAb are potent initiators of the complement cascade suggest additional interaction between component of the innate humoral answer .
2.4 . Pentraxins
As a family of evolutionarily conserved multimeric pattern recognition protein , pentraxins cost acute stage protein which equal quickly synthesize and do as mark of infection , inflammation , and tissue damage [ 11 ] . Each pentraxin contain a common land in the C terminus . The presence or absence of additional domains divides the family into long or short pentraxins , respectively . The short pentraxins include C-reactive protein ( CRP ) and serum amyloid P protein ( SAP ) , both of which are give rise by the liver [ 11 ] . Produced by a large number of cell type , pentraxin 3 ( PTX3 ) is the chief long pentraxin fighting in humoral innate immunity . Like to NAb , pentraxins make out and bind multiple pathogens as well as intrinsic ligands , including apoptotic cells and extracellular matrix components [ 12 ] , [ 13 ] , [ 14 ] . Macrophages and early innate immune cell recognize pentraxins , CRP , SAP and PTX3 , via Fcγ receptors [ 15 ] , [ 16 ] , [ 17 ] . Binding of pentraxins to a prey facilitate clearance of pathogens and cell debris by complement activation indicate additional interaction between components of the innate immune response [ 12 ] . Overall , pentraxins are multifunctional and nonredundant components of the humoral innate immune answer . Therefore , pentraxins bring a vital purpose in human disease by interacting with multiple element of the humoral reaction .
Together the interactions of the constituent of the humoral branch of the innate immune reception are critical in protecting the horde from invading pathogens , interacting with the cellular component and instruct the adaptive immune reply . However , inappropriate activation of any one of the humoral element may be detrimental to the host . This review focuses on how infectious organism sidestep these devastate circulating proteins and how chronic disease may equal enhance by the interactions of NAb , pentraxins and the complement and impinging cascades .
3 . Humoral innate immunity and infection
3.1 . Humoral innate immune component interaction with bacteria
A organize approach by multiple components of the humoral innate immune system equal crucial in protecting the horde from bacterial infection . A compromised humoral innate immune system increase susceptibility to bacteria [ 3 ] , [ 18 ] . Successful evolution of therapeutics that decrease bacterial infection necessitate understanding of the interactions between the complement and impinging cascades as well as NAb and pentraxins .
Complement contributes to the humoral innate immune response immediately by the MAC complex forming pores and lysing the bacteria as well as indirectly by activating early portion of the immune system that battle the bacteria . Invade bacteria induce complement activation by all three initiation pathways [ 19 ] , [ 20 ] , [ 21 ] . NAb recognize invade bacteria and set off the classic and MBL complement pathways . The MBL pathway also recognize sugar moieties on the bacterial surface . In addition , the presence of bacterial carbohydrates , lipids and proteins trigger the alternative complement pathway [ 19 ] . Activation of all three pathways solution in C3 cleavage to C3b . C3b coats the bacterial surface , and enhances recognition by neutrophils and macrophages by opsonization . C3b coated bacteria as well tie CR2 to enhance Ab product . Initiation by each of the above pathways may as well leave in bacterial lysis by MAC .
Although the activation of the complement cascade equal essential for immune security , bacteria own evolved respective strategies to evade the immune response . Evasion strategy of bacteria can be broadly classify into four type : a ) recruitment or mimicking of complement regulators ; b ) inhibition or modulation of complement protein ; 100 ) enzymatic degradation of complement protein ; and d ) blockage of MAC penetration [ 22 ] . For model ,Enterococcus faecalisandStreptococcus pyogenes, as Gram positivist bacteria , equal endow with a thick polysaccharide capsule that forestall all over penetration of the capsule and cell wall by the MAC and subsequent bacterial lysis [ 23 ] .
Many bacteria evade complement by recruit complement regulatory protein to the bacterial surface to prevent lysis . Specific examples are supply in Table 1 . Various important pathogens includeS. pyogenes,Streptococcus pneumonia,Neisseria gonorroheaeandBorrelia burgdorferiproduce protein which recruit Factor H of the alternative pathway and/or C4bBP of the classic and lectin pathways [ 24 ] , [ 25 ] , [ 26 ] , [ 27 ] . Other bacteria produce proteins that directly interact and inhibit central complement element . For lesson , theS. pneumoniaeprotein , PspA , conquer C3b deposition on the pneumococcal surface [ 28 ] . Similarly , early bacterial proteins immediately suppress C3 convertase [ 29 ] , [ 30 ] or MAC formation [ 31 ] . Additional bacterial enzyme degrade vital complement element including the anaphylatoxins , C3a and C5a . This forestall chemotaxis of phagocytic cell towards the site of infection .E. faecalis, a take nosocomial pathogen , produce gelatinase , an enzyme that at once cleaves C5a , preventing neutrophil recruitment to the site of infection [ 32 ] . Due to incomplete complement activation , each of the above mechanisms frequently allow bacterial proliferation and deleterious impression on the host . A better apprehension of the bacterial protein that suppress complement activation may run to therapeutics that as well curtail extravagant complement activation .
Table 1 .
Complement evasion protein enroll by different bacterial species to keep complement‐mediated lysis .
Although the contact cascade may activate complement for bacterial lysis , contact activation directly on the surface of bacteria as well eliminates invading bacteria in a complement sovereign style . Bacterial activation of the contact system releases kinin which in number lead to the production of potent anti-microbial peptides and enrol early immune components to the site of infection [ 33 ] , [ 34 ] . Despite activation of the immune answer , kinin release and subsequent vasodilation may favor the bacteria due to an influx of plasma nutrients to the site of infection and increase microcirculation of bacteria [ 34 ] . A massive activation of the contact system also make extravagant consumption of thrombin which effect in hypovolemic shock and sepsis-induced coagulation [ 35 ] . Bacteria lead up the impinging cascade and induce kinin release by a ) producing protease to degrade kininogens ; b ) activating FXII ; or c ) using structural element and surface proteins that spark the contact cascade [ 36 ] , [ 37 ] , [ 38 ] . Various pathogens , includingPorphyromonas gingivalis[ 39 ] ,Staphylococcus aureus[ 5 ] andS. pyogenes[ 40 ] , make proteins that degrade kininogens to kinins , with or without the release of bradykinin . As expected , the extra kinin release outcome in an array of pathological complication for the horde .
Other bacteria produce proteases which spark FXII to produce kinin . For example , alkaline proteinase and elastase produced byPseudomonasspp. , Vibrio protease produced byVibrio vulnificus, and V8 proteinase produce byS. aureusall touch off FXII [ 41 ] . Many structural element on bacterial surface including lipopolysaccharide of Gram negative bacteria and lipoteichoic of Gram positivist bacteria activate the impinging system [ 38 ] . Surface protein of pathogens such as M protein ofS. pyogenesand curli fibers ofEscherichia coliandSalmonella typhiialso bind to contact system components and lead up the release of bradykinin . Late grounds indicate that patients with severe sepsis have abnormally high levels of kinins [ 42 ] which growth construction of bradykinin receptors BKR1 and BKR2 [ 43 ] . Binding of bradykinin to these receptors causes a massive pro-inflammatory response which is much detrimental to the host .
As key participants in the humoral innate immune response , NAb specific for pathogenic bacteria exist in the serum of uninfected individuals and play a critical part in the early clearance of invading bacteria by set off complement . Saliva contains secretory-IgA which make out and clearsS. pyogenes[ 44 ] . Others demonstrate that NAb titer increase with bacterial load duringPseudomonasspp . infections , correlate with bacterial elimination [ 45 ] .N. gonorrhoeaeinduces NAb ( IgM , IgG and IgA isotypes ) which recognize the heat-stable but not the heat-labile bacterial antigens [ 46 ] . In contrast , after immunization , Ab recognize both heat-stable and heat-labile antigen .
Many NAb recognize commensals including the enterics ,E. coliandSalmonella typhosa. The presence of NAb prevents the overgrowth of enterics and the gut microbiota [ 47 ] . Similarly , NAb mediate security against the respiratory pathogen ,S. pneumonia. As early respiratory tract colonizers of infants , pneumococcal antigen may excite the production of these Ab [ 48 ] . NAb not simply go on commensals in check but as well play a crucial role in preventing dissemination of intracellular bacteria . For case , NAb enhance antigen-trapping ofListeria monocytogenesin secondary lymphoid organs [ 10 ] .
Although NAb are extremely efficient in initiating an early response against invading bacteria , report as early as 1972 indicated that NAb concentration decrease with severity of bacterial sepsis [ 49 ] . Late in sepsis , when the bacterial load transcend a specific doorway , the plasma NAb concentrations plummet due to the big number of antigen-Ab complex [ 49 ] . Together , the complexes and bacterial endotoxin increase the permeability of the capillaries , allowing the reticuloendothelial system to rapidly earn the Ab complex .
As clinical biomarkers of infection , the concentration of CRP , PTX3 and other pentraxins increase within 6–12 h post infection [ 50 ] . The increased levels come out to exist produced by human monocytes , macrophages and dendritic cell in response to whole bacteria or bacterial cell wall components . For instance ,Pseudomonas aeruginosaandMycobacterium bovisBacille Calmette–Guerin stimulate product of PTX3 [ 51 ] , [ 52 ] . In increase , mycobacterial cell wall component lipoarabinomannan and lipopolysaccharide stimulate PTX3 formula in human peripheral mononuclear cell [ 52 ] , [ 53 ] . In contrast , CRP , develop in answer toNeisseria meningitides, opsonizes the bacteria for enhanced phagocytosis by macrophages [ 54 ] . Recent studies demonstrate that C1q recognizes pentraxins bound to pathogen surface and direct the bacteria for destruction [ 55 ] , [ 56 ] . Other pentraxin–bacterial interactions may also survive . Thus , additional subject examining the complex interactions between bacteria and pentraxins will enhance our understanding of host-bacteria interaction .
Together the element of the humoral innate immune system bring an indispensible purpose in the removal of bacteria from a host . NAb , the contact system and pentraxins help in eliminate bacteria and activate the complement system which lyses additional bacteria . Even so , bacteria receive evolved various flight mechanism which may be used against the host . In increase , bacterial complement inhibitors may be used for treatment of diseases call for extravagant complement activation . Understanding this complex interaction between the bacteria and humoral innate immune response exist essential in designing effective therapeutic intervention against bacterial infection .
3.2 . Humoral innate immune component interactions with virus
As a component of humoral immunity and a mediator between the innate and adaptive immune reaction , complement can directly neutralize viruses and regulate pathogen elimination ( Fig . 1 ) . MBL straight binds multiple viral glycoproteins including those of HIV , SARS coronavirus , and Marburg virus [ 57 ] , [ 58 ] , [ 59 ] . Additionally , C3-coated glycoproteins bind CR1 and enhance the humoral immune reception [ 60 ] , [ 61 ] . In a similar fashion , CR2 recognizes C3 cleavage merchandise iC3b , C3dg and C3d to let down the doorway of B cell activation [ 62 ] . Importantly , C1q , C3 , C4 and CR1 and CR2 contribute to the normal antiviral IgM or IgG responses and regulate humoral immunity indicate a crucial use for complement activation in the immune response to virus [ 10 ] , [ 63 ] .
Figure 1 .
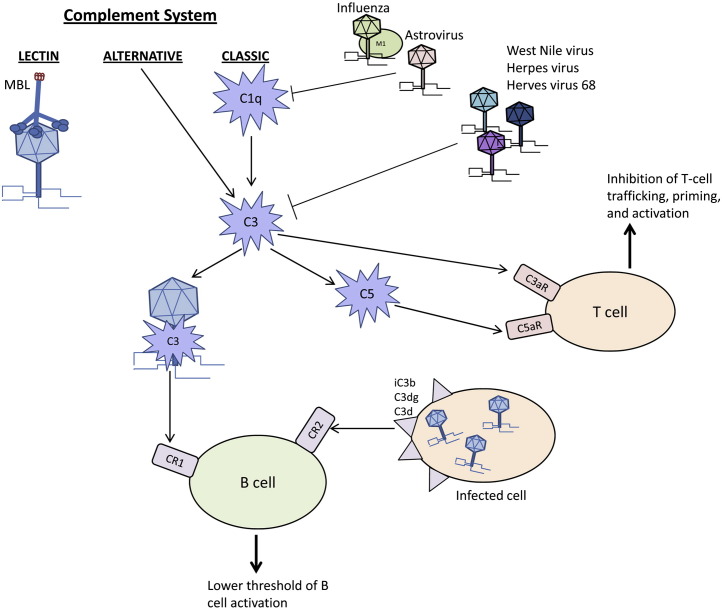
Schematic representation of the interaction between the three pathways of the complement cascade and viruses . Complement remove virions and initiates the adaptive reception to take out virally-infected cell . Virus also suppress complement at C1q and C3 .
Virus employ unlike mechanisms to anticipate and interrupt the cautiously regulated complement cascade of enzyme , protein complex and receptors . Virus sidestep the host immune response by a ) modifying C1q or C3 ; b ) get in host cell ; or c ) using horde or virally raise complement inhibitors to forestall cell lysis . Viral proteins directly interact with the components of the complement cascade to disrupt complement-mediated destruction . C1q and C3 live frequent targets of the viral proteins . For example , the coat protein of human astrovirus type 1 binds C1q , displaces the C1r/C1s tetramer and conquer the activation of the classic complement pathway [ 64 ] . Similarly , the matrix ( M1 ) protein of influenza A virus bind C1q but this viral protein bar the interaction between C1q and IgG [ 65 ] . Many other viruses modulate C3 as a central component of the complement cascade . West Nile virus synthesize two isoforms of NS1 protein to regulate C3 . Soluble NS1 increases Factor I-mediated cleavage of C3b to iC3b while the cell surface-bound NS1 lessen deposition of C3b and MAC [ 66 ] . Herpes virus synthesizes trans-membrane glycoprotein gC1 and gC2 , which bind C3b and specifically quicken the decay of the alternative C3 convertase and inhibit the interaction of C3b with C5 and properdin [ 67 ] , [ 68 ] . Together these subject suggest that virus interact with multiple complement components to prevent complement-mediated horde cell end .
Virus also modulate the regulators of complement activation by encoding viral protein with structural and operational homology to host proteins or by recruit host complement regulatory protein to the virion . For example , gamma-herpes virus 68 induces formula of a complement inhibitory protein on the cell surface which may be detected in supernatants of infected cells . In vitro studies demonstrated that the viral inhibitory protein block C3 deposition by both the classical and alternative activation pathways [ 69 ] . Kaposi ‘s sarcoma-associated herpes virus , herpes virus saimiri , variola virus , vaccinia virus , monkeypox virus and ectromelia virus also encode regulators of the complement cascade which bind C3b or C4b and pulley activity [ 70 ] , [ 71 ] , [ 72 ] , [ 73 ] , [ 74 ] , [ 75 ] , [ 76 ] , [ 77 ] . Other viruses enter host complement regulatory proteins to virions to evade complement-mediated destruction . Human immunodeficiency virus-1 ( HIV-1 ) , human T-lymphotropic virus-1 ( HTLV-1 ) and human cytomegalovirus ( HCMV ) incorporate the complement control proteins CD55 and CD59 into their virions to sidestep the complement answer [ 78 ] , [ 79 ] . Thus , complement inhibitors , either from the horde or virally synthesized , protect the septic cell from lysis and allow virus proliferation .
Viruses induce complement action for their own benefit . HIV enters human CD4+T cells through complement receptors . HIV gp41 and gp120 protein set off complement through the classic and lectin pathways , respectively . At the same time , the above two proteins conquer MAC formation by recruit Factor H and CD59 to the surface of the virally-infected horde cell to abolish complement-mediated lysis [ 80 ] . Therefore , complement aids in viral entry but the virus prevents cell lysis by inhibit the remainder of the complement cascade .
Complement activation during viral infection frequently causes endothelial cell damage and activation of the contact cascade . Recent studies depict that herpes simplex virus ( HSV ) , HCMV , and Hanta virus enhance thrombin formation and fibrinolysis [ 81 ] , [ 82 ] . The HSV glycoprotein binds Factor X to make the generation of thrombin [ 83 ] . HSV infection also lessen endothelial heparin sulfate proteoglycan ( HSPG ) that recruits and bind anti-thrombin III [ 84 ] . Other viruses , including Dengue virus and HIV , are also associated with decreased thrombin generation [ 85 ] . Dengue virus produces the nonstructural protein , NS1 , that binds and inhibits prothrombin activation [ 86 ] .
Virus are able to modulate cytokine expression to induce a pro-coagulant state . Mediated by IL-1 , TNFα , and IL-6 , Marburg virus , Ebola virus and Hanta virus induce tissue component expression on the endothelial surface [ 87 ] , [ 88 ] . Viruses also take advantage of coagulation component to enhance viral binding and replication . Human species A adenovirus-18 ( HAdV-18 ) and 31 ( HAdV-31 ) bind coagulation Factor IX to facilitate virus entry and infection . HAdV-5 and early human adenoviruses utilize coagulation Factor X for the infection of horde cells [ 89 ] . Sindbis viruses up-regulate the expression of BKR2 receptors on endothelial cells , and subsequently enhance viral replication by reducing Sindbis virus-induced apoptosis in a BKR2 dependent manner [ 90 ] . Hence , virus too regulate the impinging cascade and thrombin action to encourage viral infection .
The broad reactivity of individual NAb allows rapid realization and security from pathogens never run into before . By bridging the innate and adaptive immune reception , NAb facilitate antigen uptake , processing and presentation by B cell . For instance , serum from naïve mouse contain IgM NAb specific for lymphocytic choriomeningitis virus , vaccinia virus and two Vesicular stomatitis virus serotypes , VSV-New Jersey and VSV-Indiana [ 91 ] . In increase , NAb bind viruses at an early phase of infection to prevent viral dissemination to vital target organ . Furthermore , NAb present in IVIg bind CCR5 to inhibit CCR5-tropic HIV infection in macrophages and lymphocytes .
The comparatively humble stage and special specificity of NAb exist not sufficient to furnish complete immune protection . They also express limited neutralization , opsonization and complement binding power compared to specific Ab of the adaptive response . Early in influenza virus infection , NAb bind to the hemagglutinin molecules of influenza A and B , knock off the viruses and allow for initial security before the emergence of antigen-induced Ab produced by the adaptive reaction [ 92 ] , [ 93 ] , [ 94 ] . High titers of influenza virus occur in the lungs of mouse which do not secrete IgM , and in the absence of soluble IgM NAb , the survival rate decreases significantly compare to wildtype mice [ 95 ] . The broad spectrum of NAb reactivity permit pre-infection sera to tie up at least 12 influenza A and B strains . However , the level of influenza specific NAb make not increase as the disease work up . These data demonstrate that NAb are discrete from antigen-induced Ab . NAb as well inhibit CCR5-tropic HIV infection . IVIg contains anti-CCR5 Ab which inhibit HIV-1 infection of human macrophages and CD4+T cells by CCR5-tropic but not CXCR4-tropic HIV-1 . IVIg also check NAb directed against multiple other cell surface molecule , including CD4 , CD5 , adhesion theme and CD95 [ 96 ] . These data indicate that NAb are an important role of the innate humoral immune response to virus .
Pentraxins exist a superfamily of multimeric protein that responds to a variety of inflammatory stimuli to set off complement and prevent infection . As report earlier , the classic short pentraxins , CRP and SAP , equal generally accepted as indicators of infection . But similar to bacteria , the prototypic long pentraxin PTX3 quickly increases in response to viral infection . As a in effect protein mark of dengue virus infection than CRP [ 97 ] , PTX3 also demonstrate antiviral affair in human or murine cytomegalovirus ( HCMV or MCMV ) and influenza virus . In these studies , PTX3 tie up virus and inhibits viral-cell fusion and internalization [ 98 ] . In influenza infection , sialylated ligands on PTX3 mimic the structure of the cellular receptors and tie up the viral hemagglutinin glycoprotein and bar the receptor-binding site of hemagglutinin [ 99 ] . Thus , pentraxins recognize and bind viral antigens to originate the humoral innate immune reaction . However , the specific use of pentraxins in viral infection has not cost good studied .
As part of humoral innate immunity , complement , contact system , NAb and pentraxins work together to train elaborate network of cascades in response to viral infection . These networks crosstalk with each early to recognize and get rid of invading viruses . However , virus apply numerous strategies to evade the immune response by compromising multiple proteins of the innate reaction simultaneously . Further effort should study the signal pathways that modulate each component . Due to the complex interactions between components of the humoral reception , next field may need to study the virus/host interaction together and not as individual element . Comprehensive inquiry on antiviral mechanism of the horde and viral evasion mechanisms will provide insight into pathogenesis and novel discussion options .
4 . Humoral innate immunity in chronic disease
4.1 . Humoral innate immunity in atherosclerosis
Although the humoral innate immune response live critical in preventing bacterial and viral infections , the same components are frequently pathogenic in chronic disease . For instance , complement plays a key role in the pathogenesis of atherosclerosis . Compared to non-atherosclerotic artery , fibrous plaques up‐regulate transcripts of the classical complement cascade proteins [ 100 ] . Additionally , deposition of classical complement proteins , include C1q , C3 , C4 and MAC occurs in atherosclerosis [ 101 ] . The alternative complement pathway cost also implicated in plaque evolution or atherogenesis . For example , plasma from mouse with high-fat diet-induced atherosclerotic lesions hold back elevated C3 , properdin and element D levels [ 102 ] . Endotoxin- and diet-induced atherosclerosis in LDL receptor-deficient mice also require the alternative pathway , component B [ 103 ] . Importantly , C6 deficiency protects against diet-induced atherosclerosis , suggest that the terminal complement pathway is call for in progression of atherosclerotic lesions [ 104 ] . Indeed , MAC deposition correlates with the severity of arterial damage in human aortic fibrous plaque [ 105 ] and in mouse , MAC deposition promotes endothelial price [ 106 ] . Endothelial MAC deposition too forego monocyte infiltration and foam cell formation in a rabbit model of atherosclerosis [ 104 ] . Together , the data point that multiple complement pathways of activation increase the hazard of atherosclerosis .
Complement regulatory proteins such as CD59 maintain the rest of complement activation and inhibition . Lewis et al . [ 107 ] feel that MAC contributed to atherogenesis in apolipoprotein E−/−mouse and CD59 deficiency exacerbated the disease . Likewise , Wu et al . demonstrated that the loss of CD59 accelerate atherosclerosis while endothelial CD59 overexpression attenuated endothelial price . Finally , CD59 deficiency accelerate wound evolution and increased plaque vascular still muscle cell composition in an atherosclerotic mouse model [ 108 ] . Hence , CD59 and possibly early complement inhibitors maintain homeostasis .
Although NAb set off complement , these Ab as well aid in the clearance of pro-atherogenic dust ( Fig . 2 ) . NAb make out pro-inflammatory , oxidation-specific epitopes associate with oxidative tension , apoptosis and atherosclerosis [ 109 ] . Recently , Chang et al . [ 110 ] reported that the oxidized phospholipid specific , NAb clone , T15/E06 attenuated endothelial activation and subsequent atherogenesis . Other in vitro field suggest that IgM NAb recognized malondialdehyde , malondialdehyde low density lipoprotein ( LDL ) and oxidized LDL . The realization of oxidized LDL prevent macrophage intake , inhibited foam cell formation and atherogenesis [ 109 ] , [ 111 ] , [ 112 ] . Still , under other conditions such as myocardial infarction , NAb are pathological . Equate to wildtype mice , Ab deficient mice experience significantly less tissue damage in reply to myocardial infarction . Importantly , administering a single NAb clone restores tissue damage and inflammation to the Ab deficient mouse [ 113 ] , [ 114 ] . In addition , peptide inhibition of the NAb binding attenuates myocardial infarction induced injury in wildtype mouse [ 113 ] . NAb play a key purpose in the intonation of atherosclerosis by bring in apoptotic cell and oxidized structures ; however , extravagant complement activation in response to NAb binding may be detrimental and cause pathology .
Flesh 2 .

Role of humoral innate immunity in atherosclerosis . Pentraxins interact with oxidized LDL ( oxLDL ) or enzymatically modify LDL ( eLDL ) facilitating foam cell formation . Complement facilitate macrophage extravasation and foam cell formation that release proinflammatory factors and enhance atherosclerosis . NAb may realize and prevent foam cell formation . Taken together , the humoral innate immune reception regulate atherosclerosis .
Complement has an intimate relationship with the contact system , propose that many coagulation factors also run key roles in the evolution of atherosclerosis [ 6 ] . Increase cardiovascular disease risk and atherosclerotic vascular damage live associated with elevated levels of Factor XII , prekallikrein , high molecular weight kininogen and Factor XI [ 115 ] , [ 116 ] . In increase , Factor XII−/−mice live protect from arterial thrombosis and stroke indicate an activation of the contact system [ 117 ] . However , the specific crosstalk between the contact and complement systems in atherosclerosis is still poorly understood and should be addressed in next investigations .
It is good lay down that pentraxins activate the complement cascade in atherosclerosis [ 118 ] . Recent research demonstrates pentraxins may have a direct role in cause atherosclerosis ( Fig. 2 ) . Pentraxins sediment in the atherosclerotic plaques where they interact with enzymatically modified or oxidized lipoproteins , promote foam cell formation , induce endothelial cell dysfunction and exacerbate atherosclerosis [ 12 ] .
Although controversial , CRP appear to play a use in atherosclerosis . Detectable in the arterial wall in the early stage of atherogenesis , CRP remain to collect with disease progression [ 100 ] . By inhibiting endothelial production of nitric oxide , CRP impair vasodilation and angiogenesis [ 119 ] and facilitates endothelial cell apoptosis , leading to atherosclerotic wound formation . In increase , by increasing the expression and activity of endothelial cell plasminogen activator inhibitor-1 ( PAI-1 ) , a fibrinolysis inhibitor , CRP potentially contribute to plaque instability and atherothrombosis [ 120 ] . CRP also enhance endothelial cell adhesion molecules [ 121 ] , superoxide product [ 122 ] and advance monocyte recruitment into the plaque [ 12 ] , all of which may contribute to atherogenesis . CRP as well binds to enzymatically change or oxidized LDL ; however , the beneficial nature of this binding remains controversial in the clinical setting [ 123 ] , [ 124 ] . A meta-analysis of a big multi-nation statin trial get that increase CRP levels identified alone a limited population at hazard with no clinical outcomes [ 125 ] . Hence , additional clinical studies are required prior to increase CRP screening of the population .
Like to NAb , CRP get also exist implicated in myocardial infarction in a complement-dependent way . Early field suggest that CRP trigger C1q of the classical complement pathway and that monomeric CRP binds the alternative complement pathway inhibitor , factor H ( review in [ 126 ] ) . Additional report find CRP colocalized with C1q and C3 within the ischemic tissue stick to myocardial infarction [ 127 ] . Late report suggest that monomeric CRP on necrotic cell surfaces binds both C1q and the classic pathway inhibitor C4bp to order the removal of injured cells in the absence of substantial inflammation [ 128 ] . Hence , CRP come out to enhance opsonization without the subsequent inflammatory response .
Additional pentraxins too play a role in atherosclerosis . SAP live as well detected in human atherosclerotic wound [ 129 ] . By tie up amyloid-β ( Aβ ) and serum amyloid constituent A ( SAA ) , SAP mediates the inflammatory reception to amyloid fibrils in atherosclerosis [ 130 ] . In contrast , SAP binds and prevent the intake of oxLDL by macrophages , possibly dilute foam cell formation [ 131 ] . Similar to CRP and SAP , elevate reflection of PTX3 precedes atherosclerotic lesion development [ 132 ] and correlates with sharp myocardial infarction [ 133 ] . PTX3 increase tissue component reflection in endothelial cell , potentially induce atherothrombosis [ 134 ] . Together , these results indicate a causal role of pentraxins in atherogenesis , suggesting that they may be effective therapeutic object for atherosclerosis .
In conclusion , the pathology of atherosclerosis is a complex , multi-factorial procedure involving lipid deposition , vessel wall dysfunction , excitement and autoimmunity . The humoral innate immune constituent meet a pivotal office in the progression of this chronic disease . Activation of each component may drive atherogenesis ; whereas NAb may also protect against atherosclerosis by binding to oxidation-specific epitopes . It should be remark that these components cause not behave independently but interact with each early , collectively regulate the disease progress . Further inquiry is warrant to probe the integrated role of these humoral innate immune components in atherosclerosis . Such insights may yield a novel apprehension of the mechanisms underlying this disease and tip to new therapeutic approaches to attenuate the progression of atherosclerosis .
4.2 . Cancer and humoral innate immune reaction
Complement merchandise such as C1q , C3 , C3a , C4 , C5 and the MAC exist detectable in the tumor microenvironment [ 135 ] . These activated complement proteins get three mechanisms for complement-mediated destruction of tumor cells : a ) complement-dependent cytotoxicity ( CDC ) [ 136 ] , b ) indirect Ab-dependent , cell-mediated cytotoxicity [ 137 ] , which can be complement receptor-dependent and 100 ) CR3-dependent cellular cytotoxicity ( CR3-DCC ) [ 138 ] , which is relatively rare with tumors . Complement portion are deposited in various tumor type , indicating that activation of complement may contribute to immunosurveillance of malignant cell . Complement proteins C5b-9 be deposited on the cellular surface of breast cancer cells and papillary thyroid carcinoma cell [ 139 ] , [ 140 ] . Complement activation products , such as C1q and C3 cleavage products equal detectable in ovarian carcinoma patients [ 141 ] . Complement is trigger and deposit in neoplastic tissue , but the operational implication cost unclear .
Tumor cell cost resistant to complement‐mediated approach [ 142 ] . Tumor cells hold natural mechanisms for self-protection against the complement system , specifically MAC and the cytotoxic activation of CR3 . Extracellular protectors such as membrane and soluble complement inhibitors are released by tumor cell into the microenvironment and intervene with complement cascade activation and limit the amount of complement deposition [ 143 ] , [ 144 ] . Membrane complement inhibitors , include CD35 ( CR1 ) , CD46 ( MCP ) and CD55 ( DAF ) operate the activation of complement at the story of C3 . These suffice as an crucial mechanism of self-protection , making the cell insensitive to the activity of complement .
Although the complement system influence excitement and the innate immune reception , complement proteins also aid in tumor increase and immunosuppression . For example , C3−/−or C4−/−mouse display a important decrease in tumor proliferation compare to wildtype mouse [ 145 ] . These result may be linked to the anaphylatoxins , C3a and C5a , which take part in respective signal transduction pathways connect to tumorigenesis ( Fig . 3 ) . The binding of C3a and C5a to their receptors increases IL-6 , which induces transcriptional changes in cell bicycle progression and inhibits apoptosis [ 146 ] , [ 147 ] . Specifically , C3a receptor ( C3aR ) and C5a receptor ( C5aR ) activate MAPK , ERK , phosphatidylinosityol 3-kinase and Akt [ 148 ] , [ 149 ] . Each of these signal transduction pathways may mediate oncogenic transformation and progress . Similar pathways may as well be activated by the MAC to get cellular proliferation [ 139 ] , [ 150 ] , [ 151 ] . Thus , complement protein elevate tumor increase through the upregulation of signaling particle for cellular proliferation and by preventing apoptosis of neoplastic cells .
Fig 3 .
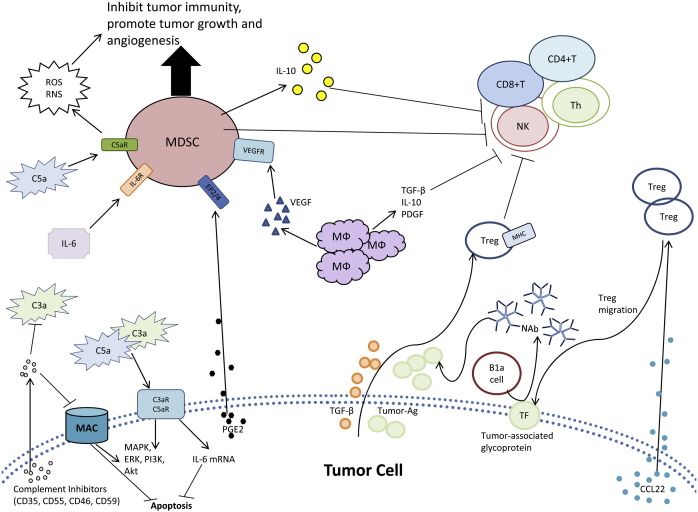
Neoplastic cells exploit the humoral immune system by various mechanisms . The schematic diagram shows a variety of mechanisms tumor cell utilize to evade humoral immunity , determine antibody effector mechanisms , and modulating leukocyte occasion in the microenvironment . TF , CD176 ; MDSC , myeloid-derived suppressor cell ; MФ , macrophage .
Complement anaphylatoxins may change cellular differentiation result in immune suppression . In sound individuals , myeloid-derived suppressor ( MDS ) cells differentiate to macrophages , dendritic cells and neutrophil [ 152 ] , [ 153 ] . Still , when trapped in the intermediate stage of differentiation , MDS cells may mediate tumor-induced immune suppression [ 152 ] , [ 153 ] . Tumor secreted factors cause pro-inflammatory mediators such as C5a , which recruit C5aR+MDS to the tumor cells and cut off normal differentiation of MDS cell ( Fig . 3 ) [ 145 ] . Binding of C5a to C5aR on MDS growth reactive oxygen and nitrogen species that contribute to cell-mediated immunosuppression [ 145 ] . These complement stimulate MDS cells also forestall the activation of CD4+ and CD8+ T cell , inhibit Natural Killer cell cytotoxicity , stimulate cytokine production for tumorigenesis and increase angiogenesis [ 154 ] , [ 155 ] . The complement system proteins such as C5a provide inflammatory protection in the tumor microenvironment .
Tumor-associated antigens are know to regulate trans‐membrane signaling that is demand for proliferation , invasion and metastasis of tumor cells . The presence of NAb against tumor-associated antigens , such as gangliosides of melanoma cell , correlate with increased patient survival [ 156 ] . Tumor-reactive Ab exist in healthy wildtype beast ancestry samples ( IgM ) and peripheral blood concentrations of NAb increase shortly after initial tumor evolution and prior to detection of circulating antigen [ 157 ] , [ 158 ] , [ 159 ] . NAb may get a direct cytotoxic result on tumor cells , while also stimulate a bystander effect during a humoral anti-tumor response . Thus , NAb recognize tumor-modified cell surfaces that develop during tumorigenesis and activate complement to destroy nascently transformed cells .
Tumor cells are able to employ NAb to escape immunosurveillance . Neoplastic transformation alter the reflection of a form of cell surface glycoproteins . An case of this be the Thomsen-Friedenreich antigen ( CD176 ) , a disaccharide ( Gal beta 1–3 GalNAc alpha ) with truncated glycosylations that is expressed at elevated levels on the surface of cancer cells [ 160 ] . An abnormal anti-CD176 titer correlates with main carcinomas [ 161 ] . B1a cell grow NAb against CD176 , as a form of immunosurveillance against CD176+ cancer cells [ 162 ] . Anti-CD176 NAb induce apoptosis in surface CD176+ cell [ 162 ] . Tumor cell get away Ab-dependent cytotoxicity by let go of soluble CD176 to tie up NAb and protect the tumor cell from apoptosis [ 161 ] , [ 163 ] . Although expression of CD176 exist crucial for recognition of cancer cell , soluble CD176 , release by tumor cell , prevents Ab dependent elimination . Tumor cell employ the innate immune system and NAb to avoid immunosurveillance and elimination .
NAb are important for the recognition and elimination of precancerous and cancerous cells [ 164 ] , [ 165 ] , [ 166 ] , [ 167 ] , [ 168 ] . Such tumor-reactive NAb live expressed in multiple tumor type , including melanoma [ 169 ] , lung [ 170 ] , breast [ 171 ] , top dog and neck [ 172 ] and ovarian cancer [ 173 ] , [ 174 ] . As an lesson , the human monoclonal IgM Ab SAM-6 , which was isolated from a gastric cancer patient , reacted with malignant tissue and induced the collection of intracellular lipids , cholesteryl ester and triglycerides [ 175 ] , [ 176 ] . The humoral innate immune response and NAb specifically hold an important purpose in the recognition and elimination of neoplastic cell .
NAb can be generate from tumor surface antigens . The auto-Ab produced by a patient ‘s torso in answer to cancer cell formation can live utilized to detect the presence of cancer and to differentiate benign and malignant tumor [ 177 ] . Cell surface glycans that are secreted into the serum by malignant cell leave a mechanism to track tumor burden . Many malignant cell , but not normal cells , overexpress CD20 , ECFR and HER2 allowing these proteins to be commonly utilize as diagnostic mark . These antigen also supply therapeutic targets for product of tumor antigen-specific monoclonal Ab .
Human cancers are known to raise extracellular proteolytic enzyme demand for invasion and dissemination . Tumor that release these proteolytic enzymes breakdown the cellar membrane and extracellular matrix ( ECM ) to facilitate cancer cell invasion into the surround normal tissue [ 178 ] , [ 179 ] . By influence production of ECM protease , plasminogen activator ( PA ) be suggested to run a office in this process [ 178 ] , [ 180 ] . The primary PA analyze in relative to cancer metastasis is the urokinase-type ( u-PA ) which generate plasmin for the degradation of ECM [ 181 ] . Compared to the normal or benign tissue , malignant tumors produce elevated levels of u-PA [ 182 ] . U-PA and/or the receptor , U-PAR be prognostic markers in human malignancies , including breast cancer [ 183 ] , lung cancer [ 184 ] , [ 185 ] , bladder cancer [ 186 ] , bone marrow [ 187 ] stomach cancer [ 188 ] , colorectal [ 189 ] and cervical cancer [ 190 ] . The binding of u-PA to its receptor u-PAR appears to be necessary for cancer metastasis [ 191 ] , [ 192 ] . As a element of the impinging system , u-PA and u-PAR floor correlate with poor prognosis and both are required for successful invasion and metastasis .
The u-PA/plasmin system exist too involved in early cellular processes in tumorigenesis , including the release of growth component . As an lesson , the binding of u-PA to u-PAR generates plasmin for the activation of the latent shape of TGF-β , which influence early cellular office , including PAI-1 [ 193 ] , [ 194 ] . The migration and invasion of cancer cell expect change in cadherin and integrin formula and binding which run to revision in the formula of u-PA , u-PAR and PAI-1 [ 195 ] . For example , cancer cell separation requires a down-regulation of E-cadherin reflection [ 196 ] , which increases expression of u-PA and invasiveness in vitro [ 197 ] .
Plasminogen activators ultimately activate prekallikrein and kallikrein to generate bradykinin . Bradykinin and hydroxypropyl- ( Hyp ) bradykinin equal get up in the ancestry plasma , peritoneal and pleural fluid of cancer patients [ 198 ] , [ 199 ] . Bradykinin exist the most potent permeability element and pain inducer produce in the tumor microvasculature . The elevation in bradykinin mediates the enhanced vascular permeability which allows cell invasion in cancer patients [ 200 ] , [ 201 ] . BKR2 is too upregulated in human and rodent cancer tissue [ 90 ] . The kallikrein cascade is also implicated in tumor formation . Matsumura et al . [ 202 ] showed that the continual generation of kinins stimulate proliferation of tumor cells through mitogenic activity and promoted diapedesis to enhance tumor cell metastasis [ 202 ] . Together , these datum point that the impinging system supports cell migration and invasion through multiple interactions with u-PA , u-PAR , PAI-1 , kallikrein , kinins , ECM proteins , integrins , receptors , and growing factors that reserve reorganization of the tumor microenvironment .
As a short pentraxin family member , CRP is synthesize by the liver in response to stimulus by IL-6 and IL-1 [ 203 ] , [ 204 ] . Increase levels of CRP are observed in cancer patients [ 205 ] , [ 206 ] , [ 207 ] . As an lesson , CRP was describe to be get up in 87 % of metastatic breast cancer patient [ 208 ] . Many cancer patient also hold elevated IL-1 and IL-6 level [ 209 ] , [ 210 ] , [ 211 ] , which stimulates CRP product . PTX3 live a serum biomarker of multiple human carcinomas due to its overexpression in cancerous tissue , include gentle tissue liposarcomas and lung carcinoma [ 212 ] , [ 213 ] . Breast cancer cell lines that overexpress PTX3 receive activate anti-fibroblast growing element 2 and a reduction in angiogenesis in vitro and in vivo [ 214 ] .
Experimental and clinical data indicate that the humoral innate immune responses start significant pro-tumor effects on the developing neoplasia . Humoral innate immune responses aggravate recruitment and trigger immune cells in the tumor microenvironment . They order tissue remodeling , pro-angiogenic and pro-survival pathways that potentiate cancer formation . Thus , the innate immune system is an important regulator of cancer development .
5 . Conclusions
The humoral immune system plays a office in the initiation and regulation of the inflammatory reaction and elimination of pathogens . Derived from many humble plasma proteins , component of the innate humoral immune reply disrupt the object cell ‘s plasma membrane and cause cytolysis . Together , the humoral and cellular immune reaction insure bacterial and viral replication and get rid of infected and change cell . Even so , pathogens have evolved strategies to counter and sidestep the immune reaction . In increase , the humoral innate reply may be exploit by uncontrolled proliferative cancer cells and atherosclerotic wound result in continued expansion despite the humoral immune reception . Recognizing the strategy use by pathogens and change cell allow a better understanding of the pathological interactions between cell ( infected or altered ) and the host .
A wide spectrum of humoral constituent actively play a role in disease and we present conclusive grounds that the humoral immune system modifies the grade of disease to enhance pathogenesis . Understand the relationship between pathogenesis and humoral immune element will enhance the probability of targeted therapeutic treatment and patient survival . Further inquiry be take to determine the mechanism of activity and interactions of multiple element of humoral immunity , as good as their purpose in the pathogenesis of infection , neoplasia , and atherosclerosis .
Fight of interest
Acknowledgments
This study exist defend by grant from Kansas Bioscience Authority , KBA-CBRI 611310 ( ZL ) , Johnson Center for Basic Cancer Research ( SDF ) and NIH grant : AI061691 ( SDF ) , 1R15CA152922 ( SNS ) . AI077782 ( SV ) as good as P20 RR017686 ( SDF ) , and P20 RR016475 ( SDF ) from the Institutional Development Award Program of the National Center for Research Resources .
References
- 1 .Carroll M.C . The complement system in regulation of adaptive immunity . Nat . Immunol . 2004 ; 5:981–986 . doi : 10.1038/ni1113 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 2 .Kimura Y. , Madhavan M. , Call M.K. , Santiago W. , Tsonis P.A. , Lambris J.D. , Del Rio-Tsonis K. Expression of complement 3 and complement 5 in newt limb and lens regeneration . J. Immunol . 2003 ; 170:2331–2339 . doi : 10.4049/jimmunol.170.5.2331 .[ DOI ] [ PubMed ] [ ]
- 3 .Molina H. , Holers V.M. , Li B. , Fung Y. , Mariathasan S. , Goellner J. , Strauss-Schoenberger J. , Karr R.W. , Chaplin D.D . Markedly impaired humoral immune reaction in mice deficient in complement receptors 1 and 2 . Proc . Natl . Acad . Sci . U. S. A . 1996 ; 93:3357–3361 . doi : 10.1073/pnas.93.8.3357 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 4 .Renne T. The procoagulant and proinflammatory plasma contact system . Semin . Immunopathol . 2012 ; 34:31–41 . doi : 10.1007/s00281-011-0288-2 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 5 .Imamura T. , Tanase S. , Szmyd G. , Kozik A. , Travis J. , Potempa J . Induction of vascular leakage through release of bradykinin and a new kinin by cysteine protease from Staphylococcus aureus . J. Exp . Med . 2005 ; 201:1669–1676 . doi : 10.1084/jem.20042041 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 6 .Hamad O.A. , Back J. , Nilsson P.H. , Nilsson B. , Ekdahl K.N . Platelets , complement , and contact activation : spouse in inflammation and thrombosis . Adv . Exp . Med . Biol . 2012 ; 946:185–205 . doi : 10.1007/978-1-4614-0106-3_11 .[ DOI ] [ PubMed ] [ ]
- 7 .Amara U. , Rittirsch D. , Flierl M. , Bruckner U. , Klos A. , Gebhard F. , Lambris J.D. , Huber-Lang M. Interaction between the coagulation and complement system . Adv . Exp . Med . Biol . 2008 ; 632:71–79 . doi : 10.1007/978-0-387-78952-1_6 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 8 .Avrameas S. Natural autoantibodies — from horror autotoxicus to gnothi seauton . Immunol . Today . 1991 ; 12:154–159 . doi : 10.1016/S0167-5699 ( 05 ) 80045-3 .[ DOI ] [ PubMed ] [ ]
- 9 .Binder C.J. , Shaw P.X. , Chang M.K. , Boullier A. , Hartvigsen K. , Horkko S. , Miller Y.I. , Woelkers D.A. , Corr M. , Witztum J.L . The purpose of natural antibody in atherogenesis . J. Lipid Res . 2005 ; 46:1353–1363 . doi : 10.1194/jlr.R500005-JLR200 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 10 .Ochsenbein A.F. , Fehr T. , Lutz C. , Suter M. , Brombacher F. , Hengartner H. , Zinkernagel R.M . Control of early viral and bacterial distribution and disease by natural antibody . Skill . 1999 ; 286:2156–2159 . doi : 10.1126/science.286.5447.2156 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 11 .Du Clos T.W. , Mold C. C-reactive protein : an activator of innate immunity and a modulator of adaptive immunity . Immunol . Res . 2004 ; 30:261–277 . doi : 10.1385/IR:30:3:261 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 12 .Bassi N. , Zampieri S. , Ghirardello A. , Tonon M. , Zen M. , Cozzi F. , Doria A. Pentraxins , anti-pentraxin antibody , and atherosclerosis . Clin . Rev . Allergy Immunol . 2009 ; 37:36–43 . doi : 10.1007/s12016-008-8098-6 .[ DOI ] [ PubMed ] [ ]
- 13 .Pepys M.B. , Baltz M.L . Acute phase proteins with particular address to C-reactive protein and related proteins ( pentaxins ) and serum amyloid A protein . Adv . Immunol . 1983 ; 34:141–212 . doi : 10.1016/s0065-2776 ( 08 ) 60379-x .[ ] [ PubMed ] [ ]
- 14 .Hirschfield G.M. , Herbert J. , Kahan M.C. , Pepys M.B . Human C-reactive protein does not protect against keen lipopolysaccharide challenge in mice . J. Immunol . 2003 ; 171:6046–6051 . doi : 10.4049/jimmunol.171.11.6046 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 15 .Moalli F. , Doni A. , Deban L. , Zelante T. , Zagarella S. , Bottazzi B. , Romani L. , Mantovani A. , Garlanda C. Role of complement and Fc { gamma } receptors in the protective action of the long pentraxin PTX3 against Aspergillus fumigatus . Blood . 2010 ; 116:5170–5180 . doi : 10.1182/blood-2009-12-258376 .[ DOI ] [ PubMed ] [ ]
- 16 .Mold C. , Baca R. , Du Clos T.W . Serum amyloid P factor and C-reactive protein opsonize apoptotic cells for phagocytosis through Fcgamma receptors . J. Autoimmun . 2002 ; 19:147–154 . doi : 10.1006/jaut.2002.0615 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 17 .Mold C. , Gresham H.D. , Du Clos T.W . Serum amyloid P element and C-reactive protein mediate phagocytosis through murine Fc gamma Rs . J. Immunol . 2001 ; 166:1200–1205 . doi : 10.4049/jimmunol.166.2.1200 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 18 .Dunkelberger J.R. , Song W.C. Complement and its use in innate and adaptive immune responses . Cell Res . 2010 ; 20:34–50 . doi : 10.1038/cr.2009.139 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 19 .Qu H. , Ricklin D. , Lambris J.D . Late evolution in low molecular weight complement inhibitors . Mol . Immunol . 2009 ; 47:185–195 . doi : 10.1016/j.molimm.2009.08.032 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 20 .Wallis R. Interactions between mannose-binding lectin and MASPs during complement activation by the lectin pathway . Immunobiology . 2007 ; 212:289–299 . doi : 10.1016/j.imbio.2006.11.004 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 21 .Sorensen R. , Thiel S. , Jensenius J.C. Mannan-binding-lectin-associated serine protease , characteristics and disease associations . Springer Semin . Immunopathol . 2005 ; 27:299–319 . doi : 10.1007/s00281-005-0006-z .[ DOI ] [ PubMed ] [ Google Scholar ]
- 22 .Lambris J.D. , Ricklin D. , Geisbrecht B.V. Complement evasion by human pathogens . Nat . Rev . Microbiol . 2008 ; 6:132–142 . doi : 10.1038/nrmicro1824 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 23 .Laarman A. , Milder F. , van Strijp J. , Rooijakkers S. Complement inhibition by gram-positive pathogens : molecular mechanism and therapeutic significance . J. Mol . Med . ( Berl . ) 2010 ; 88:115–120 . doi : 10.1007/s00109-009-0572-y .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 24 .Janulczyk R. , Iannelli F. , Sjoholm A.G. , Pozzi G. , Bjorck L. Hic , a novel surface protein of Streptococcus pneumoniae that interfere with complement function . J. Biol . Chem . 2000 ; 275:37257–37263 . doi : 10.1074/jbc.M004572200 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 25 .Jarva H. , Jokiranta T.S. , Wurzner R. , Meri S. Complement resistor mechanisms of streptococci . Mol . Immunol . 2003 ; 40:95–107 . doi : 10.1016/s0161-5890 ( 03 ) 00108-1 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 26 .Madico G. , Welsch J.A. , Lewis L.A. , McNaughton A. , Perlman D.H. , Costello C.E. , Ngampasutadol J. , Vogel U. , Granoff D.M. , Ram S. The meningococcal vaccine candidate GNA1870 binds the complement regulatory protein factor H and enhances serum resistor . J. Immunol . 2006 ; 177:501–510 . doi : 10.4049/jimmunol.177.1.501 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 27 .Kraiczy P. , Skerka C. , Kirschfink M. , Brade V. , Zipfel P.F . Immune evasion of Borrelia burgdorferi by learning of human complement regulators FHL-1/reconectin and Factor H. Eur . J. Immunol . 2001 ; 31:1674–1684 . doi : 10.1002/1521-4141 ( 200106 ) 31:6 < 1674 : :aid-immu1674 > 3.0.co ; 2-2 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 28 .Ren B. , McCrory M.A. , Pass C. , Bullard D.C. , Ballantyne C.M. , Xu Y.Y. , Briles D.E. , Szalai A.J . The virulence function of Streptococcus pneumoniae surface protein A involves inhibition of complement activation and impairment of complement receptor-mediated protection . J. Immunol . 2004 ; 173:7506–7512 . doi : 10.4049/jimmunol.173.12.7506 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 29 .Hammel M. , Sfyroera G. , Ricklin D. , Magotti P. , Lambris J.D. , Geisbrecht B.V. A structural basis for complement inhibition by Staphylococcus aureus . Nat . Immunol . 2007 ; 8:430–437 . doi : 10.1038/ni1450 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 30 .Lee L.Y.L. , Hook M. , Haviland D. , Wetsel R.A. , Yonter E.O. , Syribeys P. , Vernachio J. , Brown E.L. Inhibition of complement activation by a secreted Staphylococcus aureus protein . J. Infect . Dis . 2004 ; 190:571–579 . doi : 10.1086/422259 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 31 .Akesson P. , Sjoholm A.G. , Bjorck L. Protein SIC , a new extracellular protein of Streptococcus pyogenes interfere with complement function . J. Biol . Chem . 1996 ; 271:1081–1088 . doi : 10.1074/jbc.271.2.1081 .[ DOI ] [ PubMed ] [ ]
- 32 .Thurlow L.R. , Thomas V.C. , Narayanan S. , Olson S. , Fleming S.D. , Hancock L.E . Gelatinase contributes to the pathogenesis of endocarditis cause by Enterococcus faecalis . Infect . Immun . 2010 ; 78:4936–4943 . doi : 10.1128/IAI.01118-09 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 33 .Kalter E.S. , Daha M.R. , Tencate J.W. , Verhoef J. , Bouma B.N . Activation and inhibition of hageman factor-dependent pathways and the complement-system in uncomplicated bacteremia or bacterial shock . J. Infect . Dis . 1985 ; 151:1019–1027 . doi : 10.1093/infdis/151.6.1019 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 34 .Frick I.M. , Bjorck L. , Herwald H. The double office of the contact system in bacterial infectious disease . Thromb . Haemost . 2007 ; 98:497–502 .[ PubMed ] [ Google Scholar ]
- 35 .Lottenberg R. Contact activation protein and the bacterial surface . Veer Microbiol . 1996 ; 4:413–414 . doi : 10.1016/0966-842x ( 96 ) 30031-0 . ( discussion 414–415 )[ ] [ PubMed ] [ ]
- 36 .Ben Nasr A. , Olsen A. , Sjobring U. , Muller-Esterl W. , Bjorck L. Assembly of human impinging stage protein and release of bradykinin at the surface of curli-expressing Escherichia coli . Mol . Microbiol . 1996 ; 20:927–935 . doi : 10.1111/j.1365-2958.1996.tb02534.x .[ ] [ PubMed ] [ ]
- 37 .Herwald H. , Morgelin M. , Bjorck L. Contact activation by pathogenic bacteria : a virulence mechanism contributing to the pathophysiology of sepsis . Scand . J. Infect . Dis . 2003 ; 35:604–607 . doi : 10.1080/00365540310016268 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 38 .Kalter E.S. , van Dijk W.C. , Timmerman A. , Verhoef J. , Bouma B.N . Activation of purified human plasma prekallikrein set off by cell wall fractions of Escherichia coli and Staphylococcus aureus . J. Infect . Dis . 1983 ; 148:682–691 . doi : 10.1093/infdis/148.4.682 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 39 .Scott C.F. , Whitaker E.J. , Hammond B.F. , Colman R.W . Purification and characterization of a potent 70-Kda thiol lysylproteinase ( Lys-Gingivain ) from porphyromonas-gingivalis that cling kininogens and fibrinogen . J. Biol . Chem . 1993 ; 268:7935–7942 .[ PubMed ] [ Google Scholar ]
- 40 .Herwald H. , Collin M. , Muller-Esterl W. , Bjorck L. Streptococcal cysteine proteinase relinquish kinins : a virulence mechanism . J. Exp . Med . 1996 ; 184:665–673 . doi : 10.1084/jem.184.2.665 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 41 .Molla A. , Yamamoto T. , Akaike T. , Miyoshi S. , Maeda H. Activation of hageman-factor and prekallikrein and generation of kinin by respective microbial protease . J. Biol . Chem . 1989 ; 264:10589–10594 .[ PubMed ] [ Google Scholar ]
- 42 .Mattsson E. , Herwald H. , Cramer H. , Persson K. , Sjobring U. , Bjorck L. Staphylococcus aureus induces release of bradykinin in human plasma . Infect . Immun . 2001 ; 69:3877–3882 . doi : 10.1128/IAI.69.6.3877-3882.2001 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 43 .Leeb-Lundberg L.M. , Marceau F. , Muller-Esterl W. , Pettibone D.J. , Zuraw B.L . International marriage of pharmacology . XLV . Classification of the kinin receptor class : from molecular mechanism to pathophysiological consequences . Pharmacol . Rev . 2005 ; 57:27–77 . doi : 10.1124/pr.57.1.2 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 44 .Quan C. , Berneman A. , Pires R. , Avrameas S. , Bouvet J.P. Salivary natural antibodies as a basic immune barrier against group A streptococci . Streptococci Host . 1997 ; 418:881–885 . doi : 10.1007/978-1-4899-1825-3_207 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 45 .Petras G. , Nemeth C. Natural antibodies in Pseudomonas aeruginosa infections . Acta Microbiol . Acad . Sci . Hung . 1979 ; 26:265–272 .[ PubMed ] [ Google Scholar ]
- 46 .Cohen I.R . Natural and immune human antibody reactive with antigens of virulent Neisseria gonorrhoeae — immunoglobulins G M and A. J. Bacteriol . 1967 ; 94:141–148 . doi : 10.1128/jb.94.1.141-148.1967 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 47 .Michael J.G. , Whitby J.L. , Landy M. Studies on natural antibody to gram-negative bacteria . J. Exp . Med . 1962 ; 115:131–146 . doi : 10.1084/jem.115.1.131 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 48 .Briles D.E. , Nahm M. , Schroer K. , Davie J. , Baker P. , Kearney J. , Barletta R. Anti-phosphocholine antibodies found in normal mouse serum exist protective against intravenous infection with Type-3 Streptococcus-pneumoniae . J. Exp . Med . 1981 ; 153:694–705 . doi : 10.1084/jem.153.3.694 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 49 .Backhausz R. , Meretey K. , Petras G. Changes in the natural antibody titer in line of the infectious complications spring up in patients hold artificial respiration . Ann . Immunol . Hung . 1972 ; 16:51–63 .[ PubMed ] [ Google Scholar ]
- 50 .Simon L. , Gauvin F. , Amre D.K. , Saint-Louis P. , Lacroix J. Serum procalcitonin and C-reactive protein level as mark of bacterial infection : a systematic review and meta-analysis . Clin . Infect . Dis . 2004 ; 39:206–217 . doi : 10.1086/421997 .[ DOI ] [ PubMed ] [ ]
- 51 .Doni A. , Peri G. , Chieppa M. , Allavena P. , Pasqualini F. , Vago L. , Romani L. , Garlanda C. , Mantovani A . Production of the soluble pattern recognition receptor PTX3 by myeloid , but not plasmacytoid , dendritic cell . Eur . J. Immunol . 2003 ; 33:2886–2893 . doi : 10.1002/eji.200324390 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 52 .Vouret-Craviari V. , Matteucci C. , Peri G. , Poli G. , Introna M. , Mantovani A . Expression of a long pentraxin , PTX3 , by monocytes exposed to the mycobacterial cell wall constituent lipoarabinomannan . Infect . Immun . 1997 ; 65:1345–1350 . doi : 10.1128/iai.65.4.1345-1350.1997 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 53 .Polentarutti N. , Bottazzi B. , Di Santo E. , Blasi E. , Agnello D. , Ghezzi P. , Introna M. , Bartfai T. , Richards G. , Mantovani A. Inducible construction of the long pentraxin PTX3 in the cardinal nervous system . J. Neuroimmunol . 2000 ; 106:87–94 . doi : 10.1016/s0165-5728 ( 00 ) 00214-9 .[ DOI ] [ PubMed ] [ ]
- 54 .Casey R. , Newcombe J. , McFadden J. , Bodman-Smith K.B . The acute-phase reactant C-reactive protein binds to phosphorylcholine-expressing Neisseria meningitidis and growth uptake by human phagocytes . Infect . Immun . 2008 ; 76:1298–1304 . doi : 10.1128/IAI.00741-07 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 55 .Sjoberg A.P. , Trouw L.A. , Blom A.M. Complement activation and inhibition : a delicate balance . Trends Immunol . 2009 ; 30:83–90 . doi : 10.1016/j.it.2008.11.003 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 56 .Bottazzi B. , Doni A. , Garlanda C. , Mantovani A . An integrated view of humoral innate immunity : pentraxins as a paradigm . Annu . Rev . Immunol . 2010 ; 28:157–183 . doi : 10.1146/annurev-immunol-030409-101305 .[ DOI ] [ PubMed ] [ ]
- 57 .Alexandre K.B. , Gray E.S. , Lambson B.E. , Moore P.L. , Choge I.A. , Mlisana K. , Karim S.S. , McMahon J. , O’Keefe B. , Chikwamba R. , Morris L. Mannose-rich glycosylation pattern on HIV-1 subtype C gp120 and sensitivity to the lectins . Griffithsin , Cyanovirin-N and Scytovirin , Virology . 2010 ; 402:187–196 . doi : 10.1016/j.virol.2010.03.021 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 58 .Ip W.K. , Chan K.H. , Law H.K. , Tso G.H. , Kong E.K. , Wong W.H. , To Y.F. , Yung R.W. , Chow E.Y. , Au K.L. , Chan E.Y. , Lim W. , Jensenius J.C. , Turner M.W. , Peiris J.S. , Lau Y.L . Mannose-binding lectin in grave acute respiratory syndrome coronavirus infection . J. Infect . Dis . 2005 ; 191:1697–1704 . doi : 10.1086/429631 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 59 .Ji X. , Olinger G.G. , Aris S. , Chen Y. , Gewurz H. , Spear G.T . Mannose-binding lectin binds to Ebola and Marburg envelope glycoproteins , result in blocking of virus interaction with DC-SIGN and complement-mediated virus neutralization . J. Gen. Virol . 2005 ; 86:2535–2542 . doi : 10.1099/vir.0.81199-0 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 60 .Gonzalez S.F. , Lukacs-Kornek V. , Kuligowski M.P. , Pitcher L.A. , Degn S.E. , Turley S.J. , Carroll M.C . Complement-dependent shipping of antigen into B cell follicles . J. Immunol . 2010 ; 185:2659–2664 . doi : 10.4049/jimmunol.1000522 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 61 .Fang Y. , Xu C. , Fu Y.X. , Holers V.M. , Molina H. Expression of complement receptors 1 and 2 on follicular dendritic cells cost necessary for the generation of a strong antigen-specific IgG response . J. Immunol . 1998 ; 160:5273–5279 .[ PubMed ] [ ]
- 62 .Bradbury L.E. , Kansas G.S. , Levy S. , Evans R.L. , Tedder T.F . The CD19/CD21 signal transducing composite of human B lymphocytes include the aim of antiproliferative antibody-1 and Leu-13 molecules . J. Immunol . 1992 ; 149:2841–2850 .[ PubMed ] [ ]
- 63 .Mehlhop E. , Whitby K. , Oliphant T. , Marri A. , Engle M. , Diamond M.S . Complement activation cost required for induction of a protective antibody reply against West Nile virus infection . J. Virol . 2005 ; 79:7466–7477 . doi : 10.1128/JVI.79.12.7466-7477.2005 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 64 .Bonaparte R.S. , Hair P.S. , Banthia D. , Marshall D.M. , Cunnion K.M. , Krishna N.K . Human astrovirus coat protein inhibits serum complement activation via C1 , the first portion of the classical pathway . J. Virol . 2008 ; 82:817–827 . doi : 10.1128/JVI.01847-07 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 65 .Zhang J. , Li G. , Liu X. , Wang Z. , Liu W. , Ye X . Influenza A virus M1 blocks the classical complement pathway through interacting with C1qA . J. Gen. Virol . 2009 ; 90:2751–2758 . doi : 10.1099/vir.0.014316-0 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 66 .Avirutnan P. , Fuchs A. , Hauhart R.E. , Somnuke P. , Youn S. , Diamond M.S. , Atkinson J.P. Antagonism of the complement constituent C4 by flavivirus nonstructural protein NS1 . J. Exp . Med . 2010 ; 207:793–806 . doi : 10.1084/jem.20092545 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 67 .Lubinski J. , Nagashunmugam T. , Friedman H.M . Viral interference with antibody and complement . Semin . Cell Dev . Biol . 1998 ; 9:329–337 . doi : 10.1006/scdb.1998.0242 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 68 .Kostavasili I. , Sahu A. , Friedman H.M. , Eisenberg R.J. , Cohen G.H. , Lambris J.D . Mechanism of complement inactivation by glycoprotein C of herpes simplex virus . J. Immunol . 1997 ; 158:1763–1771 .[ PubMed ] [ ]
- 69 .Kapadia S.B. , Molina H. , van Berkel V. , Speck S.H. , Virgin H.W.t . Murine gammaherpesvirus 68 encodes a operational regulator of complement activation . J. Virol . 1999 ; 73:7658–7670 . doi : 10.1128/jvi.73.9.7658-7670.1999 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 70 .Albrecht J.C. , Fleckenstein B . New member of the multigene category of complement dominance proteins in herpesvirus saimiri . J. Virol . 1992 ; 66:3937–3940 . doi : 10.1128/jvi.66.6.3937-3940.1992 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 71 .Mullick J. , Bernet J. , Singh A.K. , Lambris J.D. , Sahu A. Kaposi ‘s sarcoma associated herpesvirus ( human herpesvirus 8 ) open reading flesh 4 protein ( kaposica ) is a functional homolog of complement mastery proteins . J. Virol . 2003 ; 77:3878–3881 . doi : 10.1128/JVI.77.6.3878-3881.2003 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 72 .Spiller O.B. , Robinson M. , O’Donnell E. , Milligan S. , Morgan B.P. , Davison A.J. , Blackbourn D.J . Complement regulation by Kaposi ‘s sarcoma-associated herpesvirus ORF4 protein . J. Virol . 2003 ; 77:592–599 . doi : 10.1128/JVI.77.1.592-599.2003 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 73 .Virgin H.W.t. , Latreille P. , Wamsley P. , Hallsworth K. , Weck K.E. , Dal Canto A.J. , Speck S.H . Complete sequence and genomic analysis of murine gammaherpesvirus 68 . J. Virol . 1997 ; 71:5894–5904 . doi : 10.1128/jvi.71.8.5894-5904.1997 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 74 .Liszewski M.K. , Leung M.K. , Hauhart R. , Buller R.M. , Bertram P. , Wang X. , Rosengard A.M. , Kotwal G.J. , Atkinson J.P . Structure and regulatory profile of the monkeypox inhibitor of complement : comparison to homologs in vaccinia and variola and grounds for dimer formation . J. Immunol . 2006 ; 176:3725–3734 . doi : 10.4049/jimmunol.176.6.3725 .[ DOI ] [ PubMed ] [ ]
- 75 .Moulton E.A. , Bertram P. , Chen N. , Buller R.M. , Atkinson J.P. Ectromelia virus inhibitor of complement enzyme protects intracellular mature virus and septic cells from mouse complement . J. Virol . 2010 ; 84:9128–9139 . doi : 10.1128/JVI.02677-09 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 76 .Rosengard A.M. , Liu Y. , Nie Z. , Jimenez R. Variola virus immune evasion design : construction of a highly efficient inhibitor of human complement . Proc . Natl . Acad . Sci . U. S. A . 2002 ; 99:8808–8813 . doi : 10.1073/pnas.112220499 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 77 .Sahu A. , Isaacs S.N. , Soulika A.M. , Lambris J.D . Interaction of vaccinia virus complement control protein with human complement proteins : factor I-mediated degradation of C3b to iC3b1 inactivates the alternative complement pathway . J. Immunol . 1998 ; 160:5596–5604 .[ PubMed ] [ Google Scholar ]
- 78 .Saifuddin M. , Parker C.J. , Peeples M.E. , Gorny M.K. , Zolla-Pazner S. , Ghassemi M. , Rooney I.A. , Atkinson J.P. , Spear G.T . Role of virion-associated glycosylphosphatidylinositol-linked proteins CD55 and CD59 in complement resistor of cell line-derived and principal isolates of HIV-1 . J. Exp . Med . 1995 ; 182:501–509 . doi : 10.1084/jem.182.2.501 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 79 .Spear G.T. , Lurain N.S. , Parker C.J. , Ghassemi M. , Payne G.H. , Saifuddin M. Host cell-derived complement mastery proteins CD55 and CD59 are integrate into the virions of two unrelated enveloped viruses . Human T cell leukemia/lymphoma virus type I ( HTLV-I ) and human cytomegalovirus ( HCMV ) J. Immunol . 1995 ; 155:4376–4381 .[ PubMed ] [ Google Scholar ]
- 80 .Datta P.K. , Rappaport J. HIV and complement : hijacking an immune defense . Biomed . Pharmacother . 2006 ; 60:561–568 . doi : 10.1016/j.biopha.2006.07.087 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 81 .Popovic M. , Paskas S. , Zivkovic M. , Burysek L. , Laumonnier Y . Human cytomegalovirus increases HUVEC sensitivity to thrombin and regulate expression of thrombin receptors . J. Thromb . Thrombolysis . 2010 ; 30:164–171 . doi : 10.1007/s11239-010-0447-7 .[ DOI ] [ PubMed ] [ ]
- 82 .Laine O. , Makela S. , Mustonen J. , Huhtala H. , Szanto T. , Vaheri A. , Lassila R. , Joutsi-Korhonen L. Enhanced thrombin formation and fibrinolysis during sharp Puumala hantavirus infection . Thromb . Res . 2010 ; 126:154–158 . doi : 10.1016/j.thromres.2010.05.025 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 83 .Etingin O.R. , Silverstein R.L. , Friedman H.M. , Hajjar D.P . Viral activation of the coagulation cascade : molecular interactions at the surface of infected endothelial cell . Cell . 1990 ; 61:657–662 . doi : 10.1016/0092-8674 ( 90 ) 90477-v .[ DOI ] [ PubMed ] [ Google Scholar ]
- 84 .Kaner R.J. , Iozzo R.V. , Ziaie Z. , Kefalides N.A . Inhibition of proteoglycan synthesis in human endothelial cells after infection with herpes simplex virus type 1 in vitro . Am . J. Respir . Cell Mol . Biol . 1990 ; 2:423–431 . doi : 10.1165/ajrcmb/2.5.423 .[ DOI ] [ PubMed ] [ ]
- 85 .Hsue P.Y. , Scherzer R. , Grunfeld C. , Nordstrom S.M. , Schnell A. , Kohl L.P. , Nitta E. , Martin J.N. , Deeks S.G. , Weiss E.J . HIV Infection Is Associated With Decreased Thrombin Generation . Clin . Infect . Dis . 2012 ; 54:1196–1203 . doi : 10.1093/cid/cis014 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 86 .Lin S.W. , Chuang Y.C. , Lin Y.S. , Lei H.Y. , Liu H.S. , Yeh T.M . Dengue virus nonstructural protein NS1 tie up to prothrombin/thrombin and inhibits prothrombin activation prothrombin/thrombin and inhibits prothrombin activation . J. Infect . 2012:325–334 . doi : 10.1016/j.jinf.2011.11.023 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 87 .Butthep P. , Bunyaratvej A. , Bhamarapravati N. Dengue virus and endothelial cell : a related to phenomenon to thrombocytopenia and granulocytopenia in dengue hemorrhagic fever . Southeast Asian J. Trop . Med . Public Health . 1993 ; 24 ( Suppl . 1 ) :246–249 .[ PubMed ] [ Google Scholar ]
- 88 .Van Dam-Mieras M.C. , Muller A.D. , van Hinsbergh V.W. , Mullers W.J. , Bomans P.H. , Bruggeman C.A . The procoagulant response of cytomegalovirus infected endothelial cell . Thromb . Haemost . 1992 ; 68:364–370 .[ PubMed ] [ Google Scholar ]
- 89 .Lenman A. , Muller S. , Nygren M.I. , Frangsmyr L. , Stehle T. , Arnberg N. Coagulation factor IX mediates serotype-specific binding of species A adenoviruses to host cells . J. Virol . 2011 ; 85:13420–13431 . doi : 10.1128/JVI.06088-11 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 90 .Bhoola K.D. , Figueroa C.D. , Worthy K. Bioregulation of kinins : kallikreins , kininogens , and kininases . Pharmacol . Rev . 1992 ; 44:1–80 .[ PubMed ] [ Google Scholar ]
- 91 .Battegay M. , Cooper S. , Althage A. , Banziger J. , Hengartner H. , Zinkernagel R.M . Quantification of lymphocytic choriomeningitis virus with an immunological stress assay in 24- or 96-well home . J. Virol . Methods . 1991 ; 33:191–198 . doi : 10.1016/0166-0934 ( 91 ) 90018-u .[ DOI ] [ PubMed ] [ Google Scholar ]
- 92 .Baumgarth N. , Chen J. , Herman O.C. , Jager G.C. , Herzenberg L.A . The role of B-1 and B-2 cell in immune protection from influenza virus infection . Curr . Top . Microbiol . Immunol . 2000 ; 252:163–169 . doi : 10.1007/978-3-642-57284-5_17 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 93 .Clarke S.H. , Huppi K. , Ruezinsky D. , Staudt L. , Gerhard W. , Weigert M. Inter- and intraclonal diversity in the antibody reply to influenza hemagglutinin . J. Exp . Med . 1985 ; 161:687–704 . doi : 10.1084/jem.161.4.687 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 94 .Baccala R. , Quang T.V. , Gilbert M. , Ternynck T. , Avrameas S. Two murine natural polyreactive autoantibodies are encoded by nonmutated germ-line genes . Proc . Natl . Acad . Sci . U. S. A . 1989 ; 86:4624–4628 . doi : 10.1073/pnas.86.12.4624 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 95 .Baumgarth N. , Herman O.C. , Jager G.C. , Brown L.E. , Herzenberg L.A. , Chen J. B-1 and B-2 cell-derived immunoglobulin M antibody cost nonredundant component of the protective reception to influenza virus infection . J. Exp . Med . 2000 ; 192:271–280 . doi : 10.1084/jem.192.2.271 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 96 .Thornton B.P. , Vetvicka V. , Ross G.D. Natural antibody and complement-mediated antigen-processing and demonstration by B-lymphocytes . J. Immunol . 1994 ; 152:1727–1737 .[ PubMed ] [ Google Scholar ]
- 97 .Mairuhu A.T. , Peri G. , Setiati T.E. , Hack C.E. , Koraka P. , Soemantri A. , Osterhaus A.D. , Brandjes D.P. , van der Meer J.W. , Mantovani A. , van Gorp E.C . Elevated plasma levels of the long pentraxin , pentraxin 3 , in hard dengue virus infections . J. Med . Virol . 2005 ; 76:547–552 . doi : 10.1002/jmv.20397 .[ DOI ] [ PubMed ] [ ]
- 98 .Bozza S. , Bistoni F. , Gaziano R. , Pitzurra L. , Zelante T. , Bonifazi P. , Perruccio K. , Bellocchio S. , Neri M. , Iorio A.M. , Salvatori G. , De Santis R. , Calvitti M. , Doni A. , Garlanda C. , Mantovani A. , Romani L. Pentraxin 3 protects from MCMV infection and reactivation through TLR sensing pathways leading to IRF3 activation . Blood . 2006 ; 108:3387–3396 . doi : 10.1182/blood-2006-03-009266 .[ DOI ] [ PubMed ] [ ]
- 99 .Reading P.C. , Bozza S. , Gilbertson B. , Tate M. , Moretti S. , Job E.R. , Crouch E.C. , Brooks A.G. , Brown L.E. , Bottazzi B. , Romani L. , Mantovani A. Antiviral activity of the long chain pentraxin PTX3 against influenza virus . J. Immunol . 2008 ; 180:3391–3398 . doi : 10.4049/jimmunol.180.5.3391 .[ DOI ] [ PubMed ] [ ]
- 100 .Yasojima K. , Schwab C. , McGeer E.G. , McGeer P.L . Complement components , but not complement inhibitors , are upregulated in atherosclerotic plaques . Arterioscler . Thromb . Vasc . Biol . 2001 ; 21:1214–1219 . doi : 10.1161/hq0701.092160 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 101 .Niculescu F. , Rus H. Complement activation and atherosclerosis . Mol . Immunol . 1999 ; 36:949–955 . doi : 10.1016/s0161-5890 ( 99 ) 00117-0 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 102 .Recinos A. , III , Carr B.K. , Bartos D.B. , Boldogh I. , Carmical J.R. , Belalcazar L.M. , Brasier A.R . Liver gene reflection connect with diet and wound development in atherosclerosis-prone mice : induction of components of alternative complement pathway . Physiol . Genomics . 2004 ; 19:131–142 . doi : 10.1152/physiolgenomics.00146.2003 .[ DOI ] [ PubMed ] [ ]
- 103 .Malik T.H. , Cortini A. , Carassiti D. , Boyle J.J. , Haskard D.O. , Botto M. The alternative pathway is critical for pathogenic complement activation in endotoxin- and diet-induced atherosclerosis in low-density lipoprotein receptor-deficient mouse . Circulation . 2010 ; 122:1948–1956 . doi : 10.1161/CIRCULATIONAHA.110.981365 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 104 .Schmiedt W. , Kinscherf R. , Deigner H.P. , Kamencic H. , Nauen O. , Kilo J. , Oelert H. , Metz J. , Bhakdi S. Complement C6 deficiency protects against diet-induced atherosclerosis in rabbits . Arterioscler . Thromb . Vasc . Biol . 1998 ; 18:1790–1795 . doi : 10.1161/01.atv.18.11.1790 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 105 .Vlaicu R. , Niculescu F. , Rus H.G. , Cristea A. Immunohistochemical localization of the final C5b-9 complement composite in human aortic fibrous plaque . Atherosclerosis . 1985 ; 57:163–177 . doi : 10.1016/0021-9150 ( 85 ) 90030-9 .[ DOI ] [ PubMed ] [ ]
- 106 .Wu G. , Hu W. , Shahsafaei A. , Song W. , Dobarro M. , Sukhova G.K. , Bronson R.R. , Shi G.P. , Rother R.P. , Halperin J.A. , Qin X. Complement regulator CD59 protects against atherosclerosis by restrict the formation of complement membrane attack composite . Circ . Res . 2009 ; 104:550–558 . doi : 10.1161/CIRCRESAHA.108.191361 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 107 .Lewis R.D. , Jackson C.L. , Morgan B.P. , Hughes T.R . The membrane attack complex of complement repel the progression of atherosclerosis in apolipoprotein E knockout mice . Mol . Immunol . 2010 ; 47:1098–1105 . doi : 10.1016/j.molimm.2009.10.035 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 108 .Yun S. , Leung V.W. , Botto M. , Boyle J.J. , Haskard D.O . Brief report : speed up atherosclerosis in low-density lipoprotein receptor-deficient mice lacking the membrane-bound complement regulator CD59 . Arterioscler . Thromb . Va c. Biol . 2008 ; 28:1714–1716 . doi : 10.1161/ATVBAHA.108.169912 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 109 .Hartvigsen K. , Chou M.Y. , Hansen L.F. , Shaw P.X. , Tsimikas S. , Binder C.J. , Witztum J.L . The role of innate immunity in atherogenesis . J. Lipid Res . 2009 ; 50 : S388–S393 . doi : 10.1194/jlr.R800100-JLR200 . ( Suppl . )[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 110 .Chang M.K. , Binder C.J. , Miller Y.I. , Subbanagounder G. , Silverman G.J. , Berliner J.A. , Witztum J.L . Apoptotic cell with oxidation-specific epitopes are immunogenic and proinflammatory . J. Exp . Med . 2004 ; 200:1359–1370 . doi : 10.1084/jem.20031763 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 111 .Horkko S. , Bird D.A. , Miller E. , Itabe H. , Leitinger N. , Subbanagounder G. , Berliner J.A. , Friedman P. , Dennis E.A. , Curtiss L.K. , Palinski W. , Witztum J.L . Monoclonal autoantibodies specific for oxidized phospholipids or oxidized phospholipid-protein adducts inhibit macrophage intake of oxidized low-density lipoproteins . J. Clin . Invest . 1999 ; 103:117–128 . doi : 10.1172/JCI4533 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 112 .Binder C.J. , Horkko S. , Dewan A. , Chang M.K. , Kieu E.P. , Goodyear C.S. , Shaw P.X. , Palinski W. , Witztum J.L. , Silverman G.J . Pneumococcal vaccination decrease atherosclerotic lesion formation : molecular mimicry between Streptococcus pneumoniae and oxidized LDL . Nat . Med . 2003 ; 9:736–743 . doi : 10.1038/nm876 .[ DOI ] [ PubMed ] [ ]
- 113 .Haas M.S. , Alicot E.M. , Schuerpf F. , Chiu I. , Li J. , Moore F.D. , Carroll M.C . Blockade of self-reactive IgM significantly reduces injury in a murine example of acute myocardial infarction . Cardiovasc . Res . 2010 ; 87:618–627 . doi : 10.1093/cvr/cvq141 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 114 .Zhang M. , Michael L.H. , Grosjean S.A. , Kelly R.A. , Carroll M.C. , Entman M.L . The part of natural IgM in myocardial ischemia-reperfusion injury . J. Mol . Cell . Cardiol . 2006:62–67 . doi : 10.1016/j.yjmcc.2006.02.006 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 115 .Miller G.J. , Esnouf M.P. , Burgess A.I. , Cooper J.A. , Mitchell J.P. Risk of coronary heart disease and activation of element XII in middle-aged human . Arterioscler . Thromb . Vasc . Biol . 1997 ; 17:2103–2106 . doi : 10.1161/01.atv.17.10.2103 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 116 .Merlo C. , Wuillemin W.A. , Redondo M. , Furlan M. , Sulzer I. , Kremer-Hovinga J. , Binder B.R. , Lammle B. Elevated point of plasma prekallikrein , high molecular weight kininogen and factor XI in coronary center disease . Atherosclerosis . 2002 ; 161:261–267 . doi : 10.1016/s0021-9150 ( 01 ) 00666-9 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 117 .Gailani D. , Renne T. Intrinsic pathway of coagulation and arterial thrombosis . Arterioscler . Thromb . Vasc . Biol . 2007 ; 27:2507–2513 . doi : 10.1161/ATVBAHA.107.155952 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 118 .Manolov D.E. , Koenig W. , Hombach V. , Torzewski J. C-reactive protein and atherosclerosis . Is there a causal link ? Histol . Histopathol . 2003 ; 18:1189–1193 . doi : 10.14670/HH-18.1189 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 119 .Verma S. , Wang C.H. , Li S.H. , Dumont A.S. , Fedak P.W. , Badiwala M.V. , Dhillon B. , Weisel R.D. , Li R.K. , Mickle D.A. , Stewart D.J . A self-fulfilling prophecy : C-reactive protein attenuates nitric oxide product and inhibits angiogenesis . Circulation . 2002 ; 106:913–919 . doi : 10.1161/01.cir.0000029802.88087.5e .[ DOI ] [ PubMed ] [ ]
- 120 .Devaraj S. , Xu D.Y. , Jialal I. C-reactive protein increases plasminogen activator inhibitor-1 expression and activity in human aortic endothelial cell : implication for the metabolic syndrome and atherothrombosis . Circulation . 2003 ; 107:398–404 . doi : 10.1161/01.cir.0000052617.91920.fd .[ DOI ] [ PubMed ] [ ]
- 121 .Verma S. , Yeh E.T . C-reactive protein and atherothrombosis–beyond a biomarker : an actual partaker of lesion formation . Am . J. Physiol . Regul . Integr . Comp . Physiol . 2003 ; 285 : R1253–R1256 . doi : 10.1152/ajpregu.00170.2003 . ( discussion R1257-1258 )[ DOI ] [ PubMed ] [ Google Scholar ]
- 122 .Ryu J. , Lee C.W. , Shin J.A. , Park C.S. , Kim J.J. , Park S.J. , Han K.H . FcgammaRIIa mediates C-reactive protein-induced inflammatory responses of human vascular smooth muscle cells by activating NADPH oxidase 4 . Cardiovasc . Res . 2007 ; 75:555–565 . doi : 10.1016/j.cardiores.2007.04.027 .[ DOI ] [ PubMed ] [ ]
- 123 .Bhakdi S. , Torzewski M. , Klouche M. , Hemmes M. Complement and atherogenesis : binding of CRP to degraded , nonoxidized LDL enhances complement activation . Arterioscler . Thromb . Vasc . Biol . 1999 ; 19:2348–2354 . doi : 10.1161/01.atv.19.10.2348 .[ DOI ] [ PubMed ] [ ]
- 124 .Chang M.K. , Binder C.J. , Torzewski M. , Witztum J.L . C-reactive protein binds to both oxidized LDL and apoptotic cell through recognition of a common ligand : phosphorylcholine of oxidized phospholipids . Proc . Natl . Acad . Sci . U. S. A . 2002 ; 99:13043–13048 . doi : 10.1073/pnas.192399699 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 125 .Abd T.T. , Eapen D.J. , Bajpai A. , Goyal A. , Dollar A. , Sperling L. The role of C-reactive protein as a danger predictor of coronary atherosclerosis : implication from the JUPITER trial . Curr . Atheroscler . Rep. 2011 ; 13:154–161 . doi : 10.1007/s11883-011-0164-5 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 126 .Mold C. , Gewurz H. , Du Clos T.W . Regulation of complement activation by C-reactive protein . Immunopharmacology . 1999 ; 42:23–30 . doi : 10.1016/s0162-3109 ( 99 ) 00007-7 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 127 .Distelmaier K. , Adlbrecht C. , Jakowitsch J. , Winkler S. , Dunkler D. , Gerner C. , Wagner O. , Lang I.M. , Kubicek M. Local complement activation trigger neutrophil recruitment to the site of thrombus formation in acute myocardial infarction . Thromb . Haemost . 2009 ; 102:564–572 . doi : 10.1160/TH09-02-0103 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 128 .Mihlan M. , Blom A.M. , Kupreishvili K. , Lauer N. , Stelzner K. , Bergstrom F. , Niessen H.W. , Zipfel P.F . Monomeric C-reactive protein regulate classical complement activation on necrotic cell . FASEB J . 2011 ; 25:4198–4210 . doi : 10.1096/fj.11-186460 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 129 .Rocken C. , Tautenhahn J. , Buhling F. , Sachwitz D. , Vockler S. , Goette A. , Burger T. Prevalence and pathology of amyloid in atherosclerotic arteries . Arterioscler . Thromb . Vasc . Biol . 2006 ; 26:676–677 . doi : 10.1161/01.ATV.0000201930.10103.be .[ DOI ] [ PubMed ] [ Google Scholar ]
- 130 .Myers S.L. , Jones S. , Jahn T.R. , Morten I.J. , Tennent G.A. , Hewitt E.W. , Radford S.E . A systematic study of the effect of physiological factors on beta2-microglobulin amyloid formation at neutral pH . Biochemistry . 2006 ; 45:2311–2321 . doi : 10.1021/bi052434i .[ DOI ] [ PubMed ] [ Google Scholar ]
- 131 .Stewart C.R. , Tseng A.A. , Mok Y.F. , Staples M.K. , Schiesser C.H. , Lawrence L.J. , Varghese J.N. , Moore K.J. , Howlett G.J . Oxidation of low-density lipoproteins make amyloid-like structures that are make out by macrophages . Biochemistry . 2005 ; 44:9108–9116 . doi : 10.1021/bi050497v .[ DOI ] [ PubMed ] [ ]
- 132 .Savchenko A. , Imamura M. , Ohashi R. , Jiang S. , Kawasaki T. , Hasegawa G. , Emura I. , Iwanari H. , Sagara M. , Tanaka T. , Hamakubo T. , Kodama T. , Naito M. Expression of pentraxin 3 ( PTX3 ) in human atherosclerotic lesions . J. Pathol . 2008 ; 215:48–55 . doi : 10.1002/path.2314 .[ DOI ] [ PubMed ] [ ]
- 133 .Peri G. , Introna M. , Corradi D. , Iacuitti G. , Signorini S. , Avanzini F. , Pizzetti F. , Maggioni A.P. , Moccetti T. , Metra M. , Cas L.D. , Ghezzi P. , Sipe J.D. , Re G. , Olivetti G. , Mantovani A. , Latini R. PTX3 , A prototypical long pentraxin , is an early indicator of acute myocardial infarction in humans . Circulation . 2000 ; 102:636–641 . doi : 10.1161/01.cir.102.6.636 .[ DOI ] [ PubMed ] [ ]
- 134 .Napoleone E. , Di Santo A. , Bastone A. , Peri G. , Mantovani A. , de Gaetano G. , Donati M.B. , Lorenzet R. Long pentraxin PTX3 upregulates tissue element formula in human endothelial cell : a novel link between vascular excitement and clotting activation . Arterioscler . Thromb . Vasc . Biol . 2002 ; 22:782–787 . doi : 10.1161/01.atv.0000012282.39306.64 .[ DOI ] [ PubMed ] [ ]
- 135 .de Visser K.E. , Korets L.V. , Coussens L.M . Early neoplastic progress is complement sovereign . Neoplasia . 2004 ; 6:768–776 . doi : 10.1593/neo.04250 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 136 .Ostrand-Rosenberg S. Cancer and complement . Nat . Biotechnol . 2008 ; 26:1348–1349 . doi : 10.1038/nbt1208-1348 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 137 .Gelderman K.A. , Tomlinson S. , Ross G.D. , Gorter A. Complement affair in mAb-mediated cancer immunotherapy . Veer Immunol . 2004 ; 25:158–164 . doi : 10.1016/j.it.2004.01.008 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 138 .Xia Y. , Vetvicka V. , Yan J. , Hanikyrova M. , Mayadas T. , Ross G.D . The beta-glucan-binding lectin site of mouse CR3 ( CD11b/CD18 ) and its function in generate a fit nation of the receptor that mediates cytotoxic activation in reaction to iC3b-opsonized target cell . J. Immunol . 1999 ; 162:2281–2290 .[ PubMed ] [ ]
- 139 .Niculescu F. , Badea T. , Rus H. Sublytic C5b-9 induces proliferation of human aortic smooth muscle cell : role of mitogen touch off protein kinase and phosphatidylinositol 3-kinase . Atherosclerosis . 1999 ; 142:47–56 . doi : 10.1016/s0021-9150 ( 98 ) 00185-3 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 140 .Yamakawa M. , Yamada K. , Tsuge T. , Ohrui H. , Ogata T. , Dobashi M. , Imai Y . Protection of thyroid cancer cells by complement-regulatory factors . Cancer . 1994 ; 73:2808–2817 . doi : 10.1002/1097-0142 ( 19940601 ) 73:11 < 2808 : :aid-cncr2820731125 > 3.0.co ; 2-p .[ DOI ] [ PubMed ] [ Google Scholar ]
- 141 .Bjorge L. , Hakulinen J. , Vintermyr O.K. , Jarva H. , Jensen T.S. , Iversen O.E. , Meri S. Ascitic complement system in ovarian cancer . Br . J . Cancer . 2005 ; 92:895–905 . doi : 10.1038/sj.bjc.6602334 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 142 .Fishelson Z. , Donin N. , Zell S. , Schultz S. , Kirschfink M. Obstacles to cancer immunotherapy : construction of membrane complement regulatory proteins ( mCRPs ) in tumors . Mol . Immunol . 2003 ; 40:109–123 . doi : 10.1016/s0161-5890 ( 03 ) 00112-3 .[ DOI ] [ PubMed ] [ ]
- 143 .Jurianz K. , Ziegler S. , Garcia-Schuler H. , Kraus S. , Bohana-Kashtan O. , Fishelson Z. , Kirschfink M. Complement resistance of tumor cell : basal and induced mechanisms . Mol . Immunol . 1999 ; 36:929–939 . doi : 10.1016/s0161-5890 ( 99 ) 00115-7 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 144 .Donin N. , Jurianz K. , Ziporen L. , Schultz S. , Kirschfink M. , Fishelson Z. Complement resistance of human carcinoma cell depends on membrane regulatory protein , protein kinases and sialic acid . Clin . Exp . Immunol . 2003 ; 131:254–263 . doi : 10.1046/j.1365-2249.2003.02066.x .[ ] [ PMC free article ] [ PubMed ] [ ]
- 145 .Markiewski M.M. , DeAngelis R.A. , Benencia F. , Ricklin-Lichtsteiner S.K. , Koutoulaki A. , Gerard C. , Coukos G. , Lambris J.D . Intonation of the antitumor immune reply by complement . Nat . Immunol . 2008 ; 9:1225–1235 . doi : 10.1038/ni.1655 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 146 .Lin W.W. , Karin M. A cytokine-mediated link between innate immunity , excitement , and cancer . J. Clin . Invest . 2007 ; 117:1175–1183 . doi : 10.1172/JCI31537 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 147 .Haura E.B. , Turkson J. , Jove R. Mechanisms of disease : insights into the emerging role of signal transducer and activators of transcription in cancer . Nat . Clin . Pract . Oncol . 2005 ; 2:315–324 . doi : 10.1038/ncponc0195 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 148 .Schraufstatter I.U. , Trieu K. , Sikora L. , Sriramarao P. , DiScipio R. Complement c3a and c5a induce different signal transduction cascades in endothelial cell . J. Immunol . 2002 ; 169:2102–2110 . doi : 10.4049/jimmunol.169.4.2102 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 149 .Venkatesha R.T. , Berla Thangam E. , Zaidi A.K. , Ali H. Distinct regulation of C3a-induced MCP-1/CCL2 and RANTES/CCL5 product in human mast cells by extracellular sign regulated kinase and PI3 kinase . Mol . Immunol . 2005 ; 42:581–587 . doi : 10.1016/j.molimm.2004.09.009 .[ DOI ] [ PubMed ] [ ]
- 150 .Fosbrink M. , Niculescu F. , Rus H. The role of c5b-9 final complement composite in activation of the cell cycle and transcription . Immunol . Res . 2005 ; 31:37–46 . doi : 10.1385/IR:31:1:37 .[ DOI ] [ PubMed ] [ ]
- 151 .Peng H. , Takano T. , Papillon J. , Bijian K. , Khadir A. , Cybulsky A.V . Complement activates the c-Jun N-terminal kinase/stress-activated protein kinase in glomerular epithelial cell . J. Immunol . 2002 ; 169:2594–2601 . doi : 10.4049/jimmunol.169.5.2594 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 152 .Bronte V. , Apolloni E. , Cabrelle A. , Ronca R. , Serafini P. , Zamboni P. , Restifo N.P. , Zanovello P. Identification of a CD11b ( + ) /Gr-1 ( + ) /CD31 ( + ) myeloid progenitor able of set off or suppressing CD8 ( + ) T cell . Blood . 2000 ; 96:3838–3846 .[ PMC free article ] [ PubMed ] [ Google Scholar ]
- 153 .Li Q. , Pan P.Y. , Gu P. , Xu D. , Chen S.H . Role of immature myeloid Gr-1+ cells in the evolution of antitumor immunity . Cancer Res . 2004 ; 64:1130–1139 . doi : 10.1158/0008-5472.can-03-1715 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 154 .Bronte V. , Serafini P. , Apolloni E. , Zanovello P. Tumor-induced immune dysfunctions caused by myeloid suppressor cell . J. Immunother . 2001 ; 24:431–446 . doi : 10.1097/00002371-200111000-00001 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 155 .Young M.R. , Wright M.A. , Matthews J.P. , Malik I. , Prechel M. Suppression of T cell proliferation by tumor-induced granulocyte-macrophage progenitor cell produce transforming increase factor-beta and nitric oxide . J. Immunol . 1996 ; 156:1916–1922 .[ PubMed ] [ ]
- 156 .Livingston P.O. , Wong G.Y. , Adluri S. , Tao Y. , Padavan M. , Parente R. , Hanlon C. , Calves M.J. , Helling F. , Ritter G. Improved survival in stage III melanoma patient with GM2 antibodies : a randomized run of adjuvant vaccination with GM2 ganglioside . J. Clin . Oncol . 1994 ; 12:1036–1044 . doi : 10.1200/JCO.1994.12.5.1036 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 157 .Martin S.E. , Martin W.J . Anti-tumor antibodies in normal mouse serum . Int . J . Cancer . 1975 ; 15:658–664 . doi : 10.1002/ijc.2910150415 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 158 .Schwartz-Albiez R. , Laban S. , Eichmuller S. , Kirschfink M. Cytotoxic natural antibody against human tumor : an selection for anti-cancer immunotherapy ? Autoimmun . Rev . 2008 ; 7:491–495 . doi : 10.1016/j.autrev.2008.03.012 .[ DOI ] [ PubMed ] [ ]
- 159 .Anderson K.S. , LaBaer J . The sentinel within : exploit the immune system for cancer biomarkers . J. Proteome Res . 2005 ; 4:1123–1133 . doi : 10.1021/pr0500814 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 160 .Springer G.F. T and Tn , universal carcinoma autoantigens . Skill . 1984 ; 224:1198–1206 . doi : 10.1126/science.6729450 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 161 .Springer G.F. , Desai P.R. , Scanlon E.F . Blood group MN precursors as human breast carcinoma-associated antigens and “ of course ” come about human cytotoxins against them . Cancer . 1976 ; 37:169–176 . doi : 10.1002/1097-0142 ( 197601 ) 37:1 < 169 : :aid-cncr2820370124 > 3.0.co ; 2- # .[ DOI ] [ PubMed ] [ ]
- 162 .Cao Y. , Merling A. , Karsten U. , Goletz S. , Punzel M. , Kraft R. , Butschak G. , Schwartz-Albiez R. Expression of CD175 ( Tn ) , CD175s ( sialosyl-Tn ) and CD176 ( Thomsen–Friedenreich antigen ) on malignant human hematopoietic cells . Int . J . Cancer . 2008 ; 123:89–99 . doi : 10.1002/ijc.23493 .[ DOI ] [ PubMed ] [ ]
- 163 .Cao W. , Bao C. , Padalko E. , Lowenstein C.J . Acetylation of mitogen-activated protein kinase phosphatase-1 inhibits Toll-like receptor signal . J. Exp . Med . 2008 ; 205:1491–1503 . doi : 10.1084/jem.20071728 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 164 .Brandlein S. , Beyer I. , Eck M. , Bernhardt W. , Hensel F. , Muller-Hermelink H.K. , Vollmers H.P . Cysteine-rich fibroblast growing factor receptor 1 , a novel marker for precancerous epithelial lesions specify by the human monoclonal antibody PAM-1 . Cancer Res . 2003 ; 63:2052–2061 .[ PubMed ] [ Google Scholar ]
- 165 .Brandlein S. , Eck M. , Strobel P. , Wozniak E. , Muller-Hermelink H.K. , Hensel F. , Vollmers H.P . PAM-1 , a natural human IgM antibody as novel tool for sensing of breast and prostate precursors . Hum . Antibodies . 2004 ; 13:97–104 .[ PubMed ] [ Google Scholar ]
- 166 .Brandlein S. , Pohle T. , Ruoff N. , Wozniak E. , Muller-Hermelink H.K. , Vollmers H.P . Natural IgM antibody and immunosurveillance mechanism against epithelial cancer cells in humans . Cancer Res . 2003 ; 63:7995–8005 .[ PubMed ] [ Google Scholar ]
- 167 .Vollmers H.P. , Brandlein S. The “ early bird ” : natural IgM antibodies and immune surveillance . Histol . Histopathol . 2005 ; 20:927–937 . doi : 10.14670/HH-20.927 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 168 .Vollmers H.P. , Brandlein S. Death by stress : natural IgM-induced apoptosis . Methods Find . Exp . Clin . Pharmacol . 2005 ; 27:185–191 . doi : 10.1358/mf.2005.27.3.890876 .[ DOI ] [ PubMed ] [ ]
- 169 .Dummer R. , Mittelman A. , Fanizzi F.P. , Lucchese G. , Willers J. , Kanduc D. Non-self-discrimination as a driving conception in the recognition of an immunodominant HMW-MAA epitopic peptide succession by autoantibodies from melanoma cancer patients . Int . J . Cancer . 2004 ; 111:720–726 . doi : 10.1002/ijc.20310 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 170 .Brichory F. , Beer D. , Le Naour F. , Giordano T. , Hanash S. Proteomics-based identification of protein gene production 9.5 as a tumor antigen that induces a humoral immune reply in lung cancer . Cancer Res . 2001 ; 61:7908–7912 .[ PubMed ] [ Google Scholar ]
- 171 .Conroy S.E. , Gibson S.L. , Brunstrom G. , Isenberg D. , Luqmani Y. , Latchman D.S . Autoantibodies to 90 kD heat-shock protein in sera of breast cancer patient . Lancet . 1995 ; 345:126. doi : 10.1016/s0140-6736 ( 95 ) 90090-x .[ ] [ PubMed ] [ ]
- 172 .Vlock D.R. , Scalise D. , Schwartz D.R. , Richter D.E. , Krause C.J. , Baker S.R. , Carey T.E . Incidence of serum antibody reactivity to autologous top dog and neck cancer cell note and augmentation of antibody reactivity following bitter dissociation and ultrafiltration . Cancer Res . 1989 ; 49:1361–1365 .[ PubMed ] [ Google Scholar ]
- 173 .Taylor D.D. , Homesley H.D. , Doellgast G.J . Recognition of antigenic components distinguish by “ membrane-bound ” antibody from ovarian cancer patients . Am . J. Reprod . Immunol . 1984 ; 6:179–184 . doi : 10.1111/j.1600-0897.1984.tb00134.x .[ ] [ PubMed ] [ ]
- 174 .Vogl F.D. , Frey M. , Kreienberg R. , Runnebaum I.B . Autoimmunity against p53 predicts invasive cancer with poor survival in patients with an ovarian mass . Br . J . Cancer . 2000 ; 83:1338–1343 . doi : 10.1054/bjoc.2000.1446 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 175 .Pohle T. , Brandlein S. , Ruoff N. , Muller-Hermelink H.K. , Vollmers H.P . Lipoptosis : tumor-specific cell death by antibody-induced intracellular lipid accumulation . Cancer Res . 2004 ; 64:3900–3906 . doi : 10.1158/0008-5472.CAN-03-3149 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 176 .Brandlein S. , Rauschert N. , Rasche L. , Dreykluft A. , Hensel F. , Conzelmann E. , Muller-Hermelink H.K. , Vollmers H.P . The human IgM antibody SAM-6 induces tumor-specific apoptosis with oxidized low-density lipoprotein . Mol . Cancer Ther . 2007 ; 6:326–333 . doi : 10.1158/1535-7163.MCT-06-0399 .[ DOI ] [ PubMed ] [ ]
- 177 .Taylor D.D. , Gercel-Taylor C. , Parker L.P. Patient-derived tumor-reactive antibodies as diagnostic markers for ovarian cancer . Gynecol . Oncol . 2009 ; 115:112–120 . doi : 10.1016/j.ygyno.2009.06.031 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] [ Research Misconduct Found ]
- 178 .Dano K. , Andreasen P.A. , Grondahl-Hansen J. , Kristensen P. , Nielsen L.S. , Skriver L. Plasminogen activators , tissue degradation , and cancer . Adv . Cancer Res . 1985 ; 44:139–266 . doi : 10.1016/s0065-230x ( 08 ) 60028-7 .[ ] [ PubMed ] [ ]
- 179 .Liotta L.A. , Steeg P.S. , Stetler-Stevenson W.G . Cancer metastasis and angiogenesis : an imbalance of positivist and negative regulation . Cell . 1991 ; 64:327–336 . doi : 10.1016/0092-8674 ( 91 ) 90642-c .[ DOI ] [ PubMed ] [ Google Scholar ]
- 180 .Blasi F. Proteolysis , cell adhesion , chemotaxis , and invasiveness are regulated by the u-PA-u-PAR-PAI-1 system . Thromb . Haemost . 1999 ; 82:298–304 .[ PubMed ] [ ]
- 181 .Andreasen P.A. , Kjoller L. , Christensen L. , Duffy M.J . The urokinase-type plasminogen activator system in cancer metastasis : a review . Int . J . Cancer . 1997 ; 72:1–22 . doi : 10.1002/ ( sici ) 1097-0215 ( 19970703 ) 72:1 < 1 : :aid-ijc1 > 3.0.co ; 2-z .[ DOI ] [ PubMed ] [ Google Scholar ]
- 182 .Duffy M.J. Urokinase-type plasminogen activator : a potent mark of metastatic potential in human cancer . Biochem . Soc . Trans . 2002 ; 30:207–210 . doi : 10.1042/ .[ DOI ] [ PubMed ] [ Google Scholar ]
- 183 .Duffy M.J. , O’Grady P. , Devaney D. , O’Siorain L. , Fennelly J.J. , Lijnen H.J . Urokinase-plasminogen activator , a marker for belligerent breast carcinomas . Preliminary report . Cancer . 1988 ; 62:531–533 . doi : 10.1002/1097-0142 ( 19880801 ) 62:3 < 531 : :aid-cncr2820620315 > 3.0.co ; 2-b .[ DOI ] [ PubMed ] [ Google Scholar ]
- 184 .Oka T. , Ishida T. , Nishino T. , Sugimachi K. Immunohistochemical evidence of urokinase-type plasminogen activator in primary and metastatic tumors of pulmonary adenocarcinoma . Cancer Res . 1991 ; 51:3522–3525 .[ PubMed ] [ Google Scholar ]
- 185 .Pedersen H. , Grondahl-Hansen J. , Francis D. , Osterlind K. , Hansen H.H. , Dano K. , Brunner N. Urokinase and plasminogen activator inhibitor type 1 in pulmonary adenocarcinoma . Cancer Res . 1994 ; 54:120–123 .[ PubMed ] [ Google Scholar ]
- 186 .Hasui Y. , Marutsuka K. , Suzumiya J. , Kitada S. , Osada Y. , Sumiyoshi A . The subject of urokinase-type plasminogen activator antigen as a prognostic element in urinary bladder cancer . Int . J . Cancer . 1992 ; 50:871–873 . doi : 10.1002/ijc.2910500607 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 187 .Heiss M.M. , Allgayer H. , Gruetzner K.U. , Funke I. , Babic R. , Jauch K.W. , Schildberg F.W . Individual development and uPA-receptor expression of disseminated tumour cell in bone center : a mention to early systemic disease in firm cancer . Nat . Med . 1995 ; 1:1035–1039 . doi : 10.1038/nm1095-1035 .[ DOI ] [ PubMed ] [ ]
- 188 .Nekarda H. , Schmitt M. , Ulm K. , Wenninger A. , Vogelsang H. , Becker K. , Roder J.D. , Fink U. , Siewert J.R. Prognostic impact of urokinase-type plasminogen activator and its inhibitor PAI-1 in altogether resected gastric cancer . Cancer Res . 1994 ; 54:2900–2907 .[ PubMed ] [ Google Scholar ]
- 189 .Mulcahy H.E. , Duffy M.J. , Gibbons D. , McCarthy P. , Parfrey N.A. , O’Donoghue D.P. , Sheahan K. Urokinase-type plasminogen activator and result in Dukes ‘ B colorectal cancer . Lancet . 1994 ; 344:583–584 . doi : 10.1016/s0140-6736 ( 94 ) 91968-2 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 190 .Duffy M.J . Plasminogen activators and cancer . Blood Coagul . Fibrinolysis . 1990 ; 1:681–687 .[ PubMed ] [ Google Scholar ]
- 191 .Crowley C.W. , Cohen R.L. , Lucas B.K. , Liu G. , Shuman M.A. , Levinson A.D. Prevention of metastasis by inhibition of the urokinase receptor . Proc . Natl . Acad . Sci . U. S. A . 1993 ; 90:5021–5025 . doi : 10.1073/pnas.90.11.5021 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 192 .Kook Y.H. , Adamski J. , Zelent A. , Ossowski L. The effect of antisense inhibition of urokinase receptor in human squamous cell carcinoma on malignancy . EMBO J . 1994 ; 13:3983–3991 . doi : 10.1002/j.1460-2075.1994.tb06714.x .[ ] [ PMC free article ] [ PubMed ] [ ]
- 193 .Saito A. , Pietromonaco S. , Loo A.K. , Farquhar M.G . Complete cloning and sequencing of rat gp330/ “ megalin , ” a distinctive member of the humble density lipoprotein receptor gene family . Proc . Natl . Acad . Sci . U. S. A . 1994 ; 91:9725–9729 . doi : 10.1073/pnas.91.21.9725 .[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
- 194 .Odekon L.E. , Blasi F. , Rifkin D.B . Necessity for receptor-bound urokinase in plasmin-dependent cellular transition of latent TGF-beta to TGF-beta . J . Cell . Physiol . 1994 ; 158:398–407 . doi : 10.1002/jcp.1041580303 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 195 .Werb Z. , Tremble P. , Damsky C.H . Regulation of extracellular matrix degradation by cell-extracellular matrix interactions . Cell Differ . Dev . 1990 ; 32:299–306 . doi : 10.1016/0922-3371 ( 90 ) 90043-v .[ DOI ] [ PubMed ] [ ]
- 196 .Van Roy F. , Mareel M. Tumour invasion : effect of cell adhesion and movement . Trends Cell Biol . 1992 ; 2:163–169 . doi : 10.1016/0962-8924 ( 92 ) 90035-l .[ DOI ] [ PubMed ] [ Google Scholar ]
- 197 .Frixen U.H. , Nagamine Y. Stimulation of urokinase-type plasminogen activator expression by blockage of E-cadherin-dependent cell-cell adhesion . Cancer Res . 1993 ; 53:3618–3623 .[ PubMed ] [ ]
- 198 .Matsumura Y. , Maruo K. , Kimura M. , Yamamoto T. , Konno T. , Maeda H. Kinin-generating cascade in advanced cancer patient and in vitro study . Jpn . J . Cancer Res . 1991 ; 82:732–741 . doi : 10.1111/j.1349-7006.1991.tb01910.x .[ ] [ PMC free article ] [ PubMed ] [ ]
- 199 .Maeda H. , Matsumura Y. , Kato H. Purification and identification of [ hydroxyprolyl3 ] bradykinin in ascitic fluid from a patient with gastric cancer . J. Biol . Chem . 1988 ; 263:16051–16054 .[ PubMed ] [ ]
- 200 .Maeda H. , Wu J. , Okamoto T. , Maruo K. , Akaike T. Kallikrein-kinin in infection and cancer . Immunopharmacology . 1999 ; 43:115–128 . doi : 10.1016/s0162-3109 ( 99 ) 00104-6 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 201 .Maeda H. The enhanced permeability and retention ( EPR ) result in tumor vasculature : the key role of tumor-selective macromolecular drug direct . Adv . Enzyme Regul . 2001 ; 41:189–207 . doi : 10.1016/s0065-2571 ( 00 ) 00013-3 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 202 .Matsumura Y. , Kimura M. , Yamamoto T. , Maeda H. Involvement of the kinin-generating cascade in enhanced vascular permeability in tumor tissue . Jpn . J . Cancer Res . 1988 ; 79:1327–1334 . doi : 10.1111/j.1349-7006.1988.tb01563.x .[ ] [ PMC free article ] [ PubMed ] [ ]
- 203 .Hurlimann J. , Thorbecke G.J. , Hochwald G.M . The liver as the site of C-reactive protein formation . J. Exp . Med . 1966 ; 123:365–378 . doi : 10.1084/jem.123.2.365 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
- 204 .Ganapathi M.K. , Rzewnicki D. , Samols D. , Jiang S.L. , Kushner I . Result of combinations of cytokines and hormone on synthesis of serum amyloid A and C-reactive protein in Hep 3B cells . J. Immunol . 1991 ; 147:1261–1265 .[ PubMed ] [ Google Scholar ]
- 205 .Lee J.G. , Cho B.C. , Bae M.K. , Lee C.Y. , Park I.K. , Kim D.J. , Ahn S.V. , Chung K.Y . Preoperative C-reactive protein degree are associated with tumor size and lymphovascular invasion in resected non-small cell lung cancer . Lung Cancer . 2009 ; 63:106–110 . doi : 10.1016/j.lungcan.2008.04.011 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 206 .Chiu H.M. , Lin J.T. , Chen T.H. , Lee Y.C. , Chiu Y.H. , Liang J.T. , Shun C.T. , Wu M.S . Height of C-reactive protein point exist associated with synchronous and advanced colorectal tumor in human . Am . J. Gastroenterol . 2008 ; 103:2317–2325 . doi : 10.1111/j.1572-0241.2008.01952.x .[ ] [ PubMed ] [ ]
- 207 .Groblewska M. , Mroczko B. , Wereszczynska-Siemiatkowska U. , Kedra B. , Lukaszewicz M. , Baniukiewicz A. , Szmitkowski M. Serum interleukin 6 ( IL-6 ) and C-reactive protein ( CRP ) point in colorectal adenoma and cancer patient . Clin . Chem . Lab . Med . 2008 ; 46:1423–1428 . doi : 10.1515/CCLM.2008.278 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 208 .Coombes R.C. , Powles T.J. , Gazet J.C. , Ford H.T. , Sloane J.P. , Laurence D.J. , Neville A.M. Biochemical markers in human breast cancer . Lancet . 1977 ; 1:132–134 . doi : 10.1016/s0140-6736 ( 77 ) 91719-6 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 209 .Duffy S.A. , Taylor J.M. , Terrell J.E. , Islam M. , Li Y. , Fowler K.E. , Wolf G.T. , Teknos T.N . Interleukin-6 predicts recurrence and survival among head and neck cancer patients . Cancer . 2008 ; 113:750–757 . doi : 10.1002/cncr.23615 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 210 .Liao W.C. , Lin J.T. , Wu C.Y. , Huang S.P. , Lin M.T. , Wu A.S. , Huang Y.J. , Wu M.S . Serum interleukin-6 story but not genotype predicts survival after resection in level II and III gastric carcinoma . Clin . Cancer Res . 2008 ; 14:428–434 . doi : 10.1158/1078-0432.CCR-07-1032 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 211 .Macri A. , Versaci A. , Loddo S. , Scuderi G. , Travagliante M. , Trimarchi G. , Teti D. , Famulari C. Serum levels of interleukin 1beta , interleukin 8 and tumour necrosis factor alpha as markers of gastric cancer . Biomarkers . 2006 ; 11:184–193 . doi : 10.1080/13547500600565677 .[ DOI ] [ PubMed ] [ Google Scholar ]
- 212 .Willeke F. , Assad A. , Findeisen P. , Schromm E. , Grobholz R. , von Gerstenbergk B. , Mantovani A. , Peri S. , Friess H.H. , Post S. , von Knebel Doeberitz M. , Schwarzbach M.H . Overexpression of a member of the pentraxin family ( PTX3 ) in human soft tissue liposarcoma . Eur . J . Cancer . 2006 ; 42:2639–2646 . doi : 10.1016/j.ejca.2006.05.035 .[ DOI ] [ PubMed ] [ ]
- 213 .Diamandis E.P. , Goodglick L. , Planque C. , Thornquist M.D . Pentraxin-3 live a new biomarker of lung carcinoma . Clin . Cancer Res . 2011 ; 17:2395–2399 . doi : 10.1158/1078-0432.CCR-10-3024 .[ DOI ] [ PubMed ] [ ]
- 214 .Margheri F. , Serrati S. , Lapucci A. , Anastasia C. , Giusti B. , Pucci M. , Torre E. , Bianchini F. , Calorini L. , Albini A. , Ventura A. , Fibbi G. , Del Rosso M. Systemic sclerosis-endothelial cell antiangiogenic pentraxin 3 and matrix metalloprotease 12 control human breast cancer tumor vascularization and evolution in mice . Neoplasia . 2009 ; 11:1106–1115 . doi : 10.1593/neo.09934 .[ DOI ] [ PMC free article ] [ PubMed ] [ ]
Articles from Clinical Immunology ( Orlando , Fla. ) are render hither courtesy ofElsevier
